The tetanic curve can be used to distinguish optimal results from sub-optimal results. This curve usually starts to form at a frequency of 60 Hz. The key factor to obtaining good results is the ability to stimulate the muscle so that it produces its maximal force and maintains that force during tetanus. The ideal curve should have an uninterrupted, sharp, vertical upswing at the time of stimulation, followed by a flat plateau phase with minimal oscillations, and an uninterrupted, sharp vertical decrease period at termination of stimulation (Figure 4). Deviations from the ideal curve are indications that the muscle is fatigued (Figure 5D) or that the muscle is not being properly stimulated to produce maximal force (Figure 5B–C). The latter generally results from incorrect electrode placement leading to failure of maximal recruitment of muscle fibers during stimulation. A distinguishing feature that allows the researcher to determine if a non-ideal curve is the result of incorrect electrode placement or pathological changes to the muscle is whether or not the tetanic curve is complete (fused) or incomplete (unfused). An unfused, incomplete tetanic curve indicates that the electrodes are misplaced, resulting in the muscle not experiencing a maximal contraction. An example of a pathological change in the muscle can be observed as decreased maximal contraction as compared to the control, or a contractile response that more rapidly fatigues.
The three different types of peaks obtained over the course of this procedure represent different electrode and leg positions and can be seen in Figure 3. The first peaks will be around 0.4N and occur when the correct electrode placement is determined superficially on the skin (Figure 3A). The second set of peaks has higher amplitude, usually around 0.5-0.6N (Figure 3B) and occur when the electrodes pierce through the dermis. After these are obtained, the leg and foot are adjusted to maximize force production, which is achieved when the peak amplitude increases to approximately 1N or greater (Figure 3C). At this point, Instant Stim can be turned off and the sequence may begin. These guidelines ensure accurate and reproducible results and are key checkpoints throughout the protocol.
The final results can be represented in different ways depending on the information that the user extracted from the force test and the experimental design. In this protocol, the maximum force is measured across all of the frequencies of stimulation, however other data points may be important for a particular researcher or application. One example is the frequency of stimulation at which the tetanic curve begins to take shape. The data can be compared to other results obtained from a previous or later experiment on the same animal, or for comparisons between different treatment groups. Force production can be normalized by body mass to calculate isometric force and provide a more unbiased assessment of the impact of age on the maximal contraction observed. Although animals of differing body weight and age will produce various maximum forces, the shape of the tetanic curve should be consistent between all groups when the procedure is performed correctly.
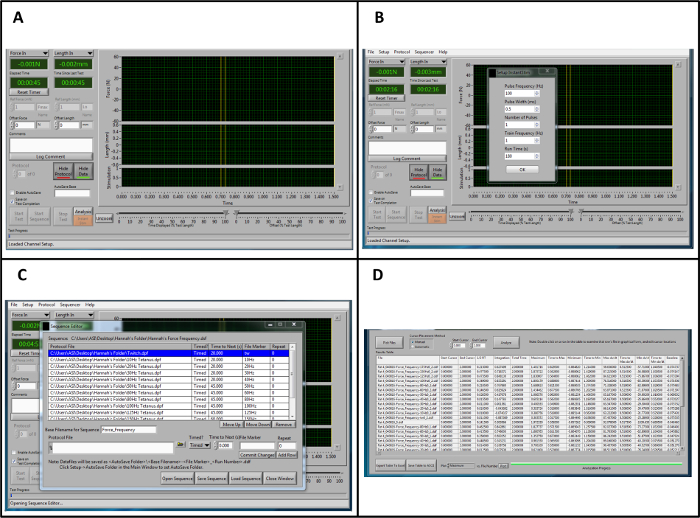
Figure 1: Overview of the Lever system Control and Data Analysis Software for Analysis. (A) Overview of the control software when opening the program. (B) Parameters for "Instant Stim." (C) Example sequence for force-frequency stimulation. (D) Representative data from a high throughput force frequency analysis in the analysis software. It should be noted that the example sequence and data analysis procedure is specific to this protocol and does not represent the full range of sequences and outputs that are provided by this software. Please click here to view a larger version of this figure.
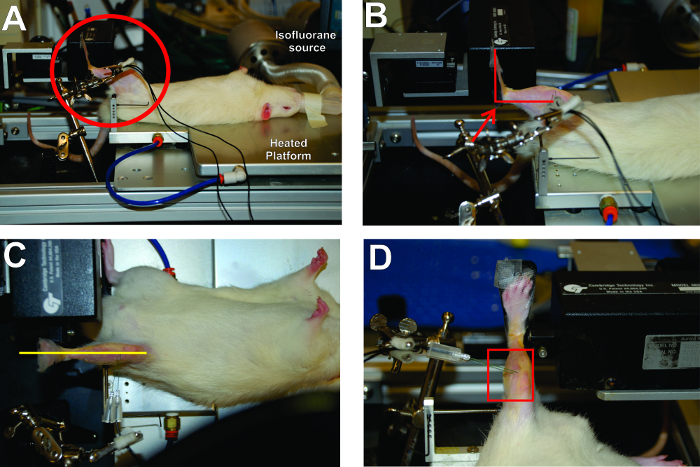
Figure 2: Critical Aspects for Positioning of the Rat and Placement of the Foot in the Apparatus. (A) The rat is in a supine position with the left foot securely attached to the footplate. The right angles made by the foot, leg, and thigh are circled. (B) The right angle created by the ankle is highlighted. (C) The leg should be aligned in a straight plane from foot to body. (D) The electrode placement is parallel and orthogonal to the plane of the peroneal nerve. Please click here to view a larger version of this figure.
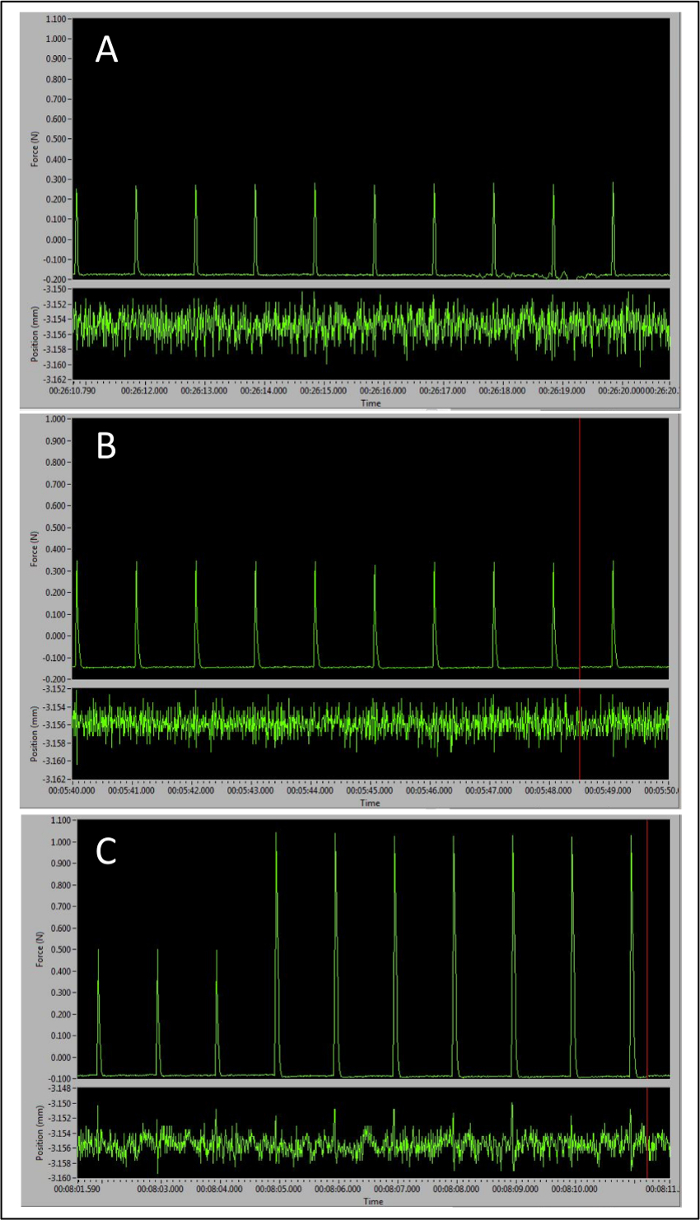
Figure 3: Representative Peaks Demonstrating the Importance of Correct Electrode Placement to Maximal Force Production. (A) Baseline peak tetanic responses observed with electrodes placed too superficially. (B) Larger peaks with electrodes inserted in the correct place. (C) Transition from larger peaks signaling correct electrode placement to optimal pre-sequence peak amplitude as the leg and foot positions are optimally adjusted. Please click here to view a larger version of this figure.
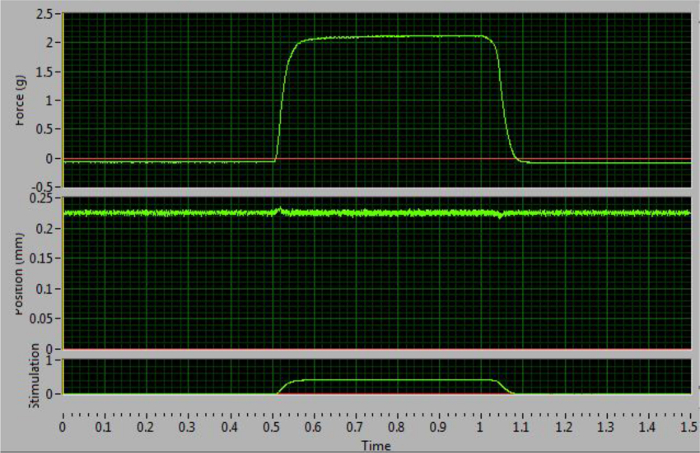
Figure 4: Optimal Tetanic Curve at 100 Hz. This curve increases and decreases sharply and has a flat plateau phase. This example indicates correct electrode placement and maximal force stimulation. Please click here to view a larger version of this figure.
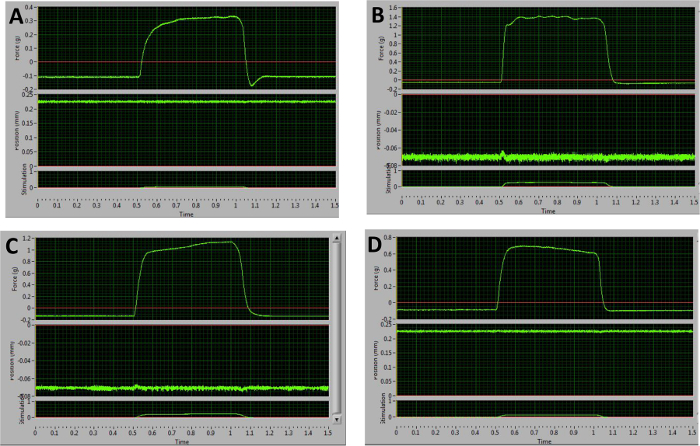
Figure 5: Representative Examples of Sub-optimal Tetanic Curves Obtained at 100 Hz. (A) Following relaxation, this curve dips below the baseline. This is indicative of stimulation of antagonists. (B–D) These graphs are the result of improper electrode placement and unequal recruitment of muscle fibers. The plateau phases demonstrate large oscillations (B), an upward slope (C), or a downward slope (D). Please click here to view a larger version of this figure.
