Different blood vessels were successfully subjected to the described isolation protocol (Figure 2). It was possible to dissect and invert aorta, vena cava, vena porta, and coronary artery from healthy dogs (all vessels from each dog, n= 4). With the same approach ECs were isolated from two congenital portosystemic shunts (extrahepatic and intrahepatic, n= 1 each). Although aorta was easily inverted, thoracic aorta segments were more challenging than abdominal aorta. In thoracic segments the aorta has many intercostal arteries branching from it, which need to be ligated individually to ensure strictly endothelial exposure to the digestion solution. The dissected segment of caudal vena cava included the branching point of the renal veins, which needed to be ligated before inversion. For the portal vein the contributing branch of the vena gastroduodenalis was ligated before inversion of the blood vessel. The coronary artery in dogs is a much smaller blood vessel (approximately 1 – 2 mm diameter in a medium sized dog), from which we excised a segment of the circumflex branch. Because it has a small diameter, it proved to be easier to invert a rather short segment of 1 cm because the Mosquito forceps could not be inserted much further.
One day post isolation adhered cells were visible in the culture plate. In culture, CaPECs had a polygonal shape and displayed a tendency to grow in patches as shown in Figure 3. Numerous colonies of endothelial cells could be observed 3 – 6 days after isolation. After approximately 10 days in culture a confluency of 80% was reached and cells could be surpassed. On average the primary endothelial cultures could be maintained for a maximum of 8 passages (once weekly at a split rate of 1:4) at which point they stopped growing.
Isolated endothelial cells expressed endothelial cell marker CD31 as indicated by qPCR (Figure 4). The expression in endothelial cells derived from aorta, vena cava, and vena porta from four dogs was compared with a control culture of CnAOECs. The cultured primary cells had a comparable CD31 expression with the control ECs (Kruskall-Wallis, p=0.856).
CaPECs derived from aorta, vena cava and vena porta showed branching after 6 hr incubation on the angiogenesis slide as shown in Figure 5.
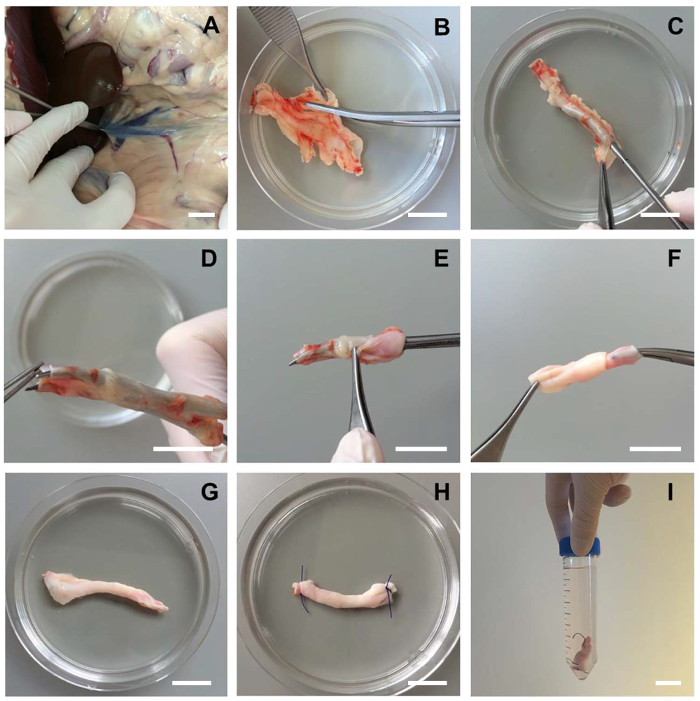
Figure 1: Dissecting and Inverting Blood Vessels for Endothelial Cell Isolation. A) The blood vessel of interest is aseptically removed from a fresh canine cadaver. B) Adherent tissue and/or fat surrounding the vessel should be removed carefully with surgical scissors without damaging the vessel itself (canine vena cava, abdominal segment of 5 cm). C) With a curved Halsted Mosquito forceps, the vessel can be entered without perforating the endothelial layer. D-G) Secure the forceps on the other end of the blood vessel and gently retract, thereby completely inverting the blood vessel. The endothelial layer is now on the outside of the vessel. H) Purse-string sutures are placed at the ends closing off the non-endothelial surface of the inverted vessel. I) The blood vessel is transferred to a 50 ml tube for washing and subsequent digestion. Scale bars indicate 2 cm. Please click here to view a larger version of this figure.
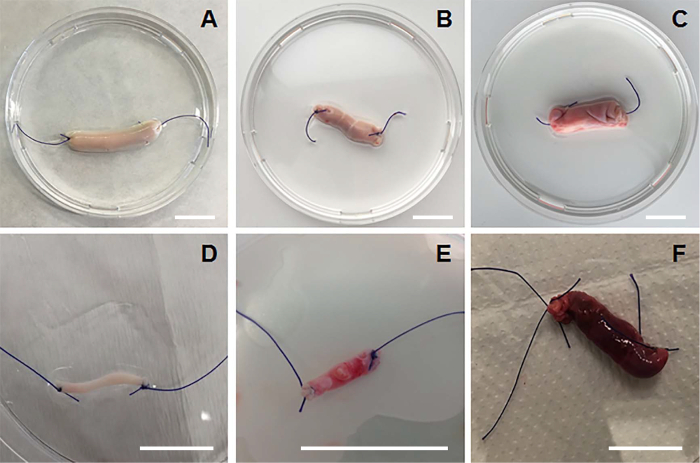
Figure 2: Inverted Vessels of Different Origin. A) An inverted aorta segment (healthy dog). B) An inverted vena porta (healthy dog). C) An inverted vena cava segment (healthy dog). D) An inverted coronary artery segment (healthy dog). E) An inverted extrahepatic portosystemic shunt derived from a Cairn terrier (age: 6 weeks old). F) An inverted intrahepatic portosystemic shunt derived from an Irish wolfhound (age: 8 weeks old). Scale bars indicate 2 cm. Please click here to view a larger version of this figure.

Figure 3: Cell Morphology. Pictures were taken two weeks after digestion of the vessels. A) ECs derived from canine aorta in passage 2. B) ECs derived from canine vena cava in passage 2. C) ECs derived from canine vena porta in passage 2. All pictures are taken with 4X original magnification. Scale bars indicate 500 µm. Insert shows 10X magnification. Please click here to view a larger version of this figure.
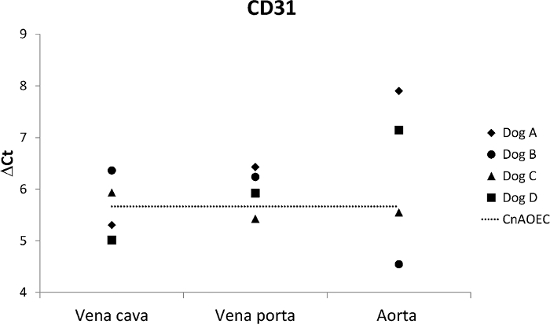
Figure 4: Gene Expression of CD31. Expression of CD31 in endothelial cells in passage 1 derived from aorta, vena cava, and vena porta (n= 4 dogs). No significant expression differences were observed between the CaPECs and the CnAOECs. Please click here to view a larger version of this figure.
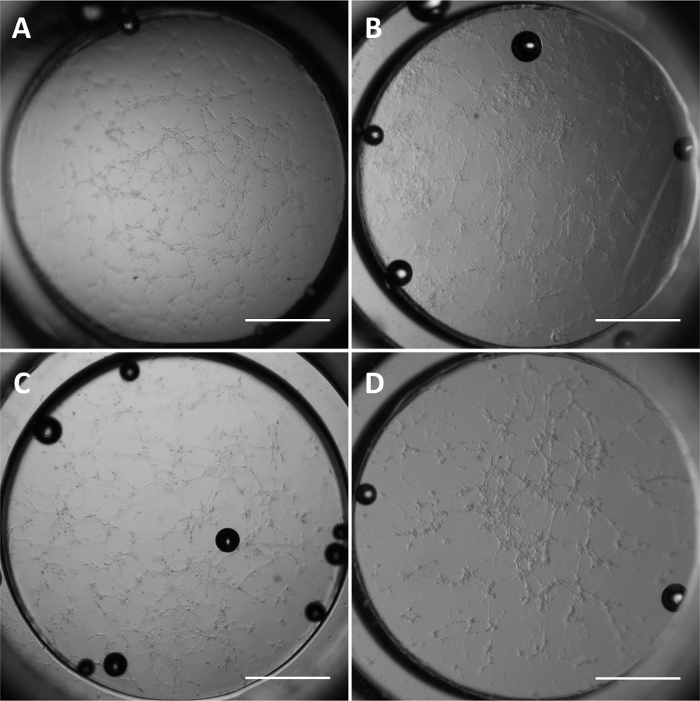
Figure 5: Angiogenesis. Photographs of CnAOECs and CaPECs from aorta, vena cava and vena porta after 6 hr incubation on an angiogenesis slide (20X magnification). Branch formation is visible after 6 hr in culture. A) CnAOECs. B) CaPECs derived from aorta. C) CaPECs derived from vena cava. D) CaPECs derived from vena porta. All cells were in passage 3. Scale bars indicate 1 mm. Please click here to view a larger version of this figure.
| GOI | Direction | 5’-Sequence-3’ | Tm annealing | Product size (bp) | Genebank number | |
| CD31 | Forward | GTTCTGCGTGTCAAGGTG | 59 °C | 85 | XM_005624261.1 | |
| Reverse | TGTCCTTCCCAAACTCCA | |||||
| beta-actin | Forward | GATATCGCTGCGCTTGTGGTC | 58 °C | 384 | NM_001195845 | |
| Reverse | GGCTGGGGTGTTGAAAGTCTC | |||||
| RPS19 | Forward | CCTTCCTCAAAAAGTCTGGG | 63 °C | 95 | XM_005616513 | |
| Reverse | GTTCTCATCGTAGGGAGCAAG | |||||
| B2MG | Forward | TCCTCATCCTCCTCGCT | 63 °C | 85 | AB745507 | |
| Reverse | TTCTCTGCTGGGTGTCG | |||||
Table 1: qPCR Primer Sets. qPCR primers for canine reference genes and CD31.
