A viral load of 1011 vg of AAV9-miR-298 was injected through a single tail-vein injection into 5 week old SBMA mice. These mice carry the human AR transgene with abnormally expanded polyglutamine tract in the AR (AR97Q) and develop signs of neuromuscular disease by 10 weeks of age (weight loss, hunched back, and muscle atrophy).11 Lumbar spinal cord and quadriceps muscle were harvested at 2, 4, 8 and 12 weeks after administration for miRNA quantification, biochemical assay and immunohistochemistry (Figure 1). Administration of the treatment and the subsequent analyses were performed by blinded investigators.
qRT-PCR analysis showed miR-298 expression in the skeletal muscle and the spinal cord two weeks and four weeks after treatment respectively, with peak expression levels at 8 weeks in the skeletal muscle and 12 weeks in the spinal cord after the injection (Figure 2). Using a microscope (Axiovert 100 M), green fluorescence signal was detected in muscle tissue and in spinal motor neurons by co-localization of GFP and the motor neuron marker choline acetyltransferase (ChAT) 10 weeks after treatment, when the mice start to show disease manifestations (Figure 2).
Using the same dose regimen, a cohort of SBMA mice was randomized to receive either miR-298 or mock at 7 weeks of age via tail vein injection for biochemical analyses and functional characterization. Injection was followed by weekly weight and behavioral assessment up to 40 weeks of age. qRT-PCR analysis showed that miR-298 treatment reduces the levels of mutant AR in affected tissues (Figure 3), and increased body weight and improved motor performance (Figure 4) starting at 10 weeks after the injection.
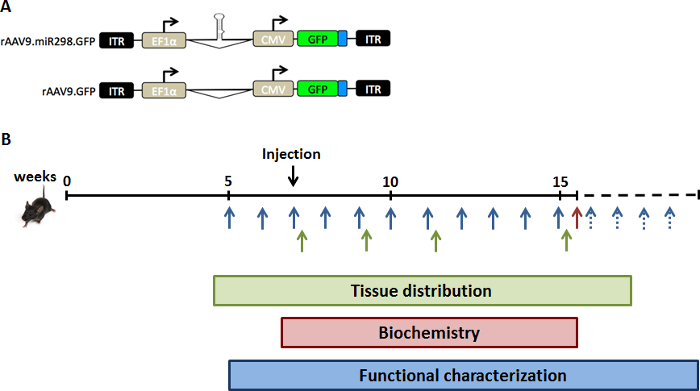
Figure 1: Schematic of study design. To increase expression levels of miR-298 in vivo, we injected SBMA mice with (A) the dual promoter AAV vector plasmid expressing GFP and either miR-298 or mock. (B) Mice were injected via a single tail-vain injection at 7 weeks of age (pre-symptomatic stage). Spinal cord and quadriceps muscle were collected from a cohort of SBMA mice at different time points for tissue distribution analysis (green). A cohort of SBMA mice was treated and sacrificed at 16 weeks of age for biochemical analyses (red). Weight and behavioral assays were performed weekly up to 40 weeks of age for functional characterization (blue). Please click here to view a larger version of this figure.
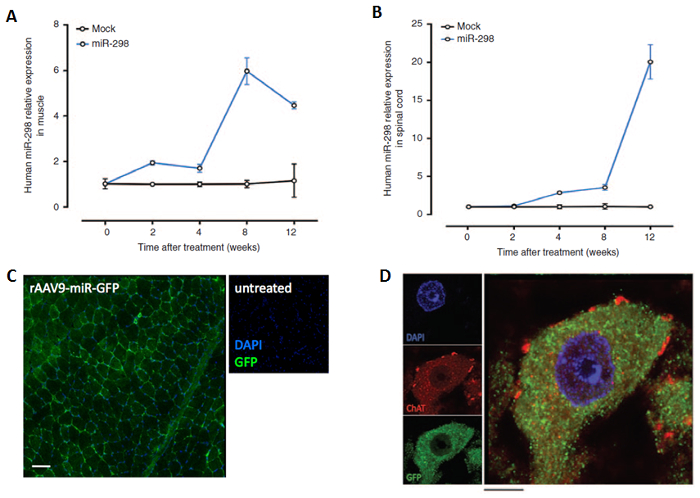
Figure 2: AAV9-miR-298 delivery in mice. Using the method described here, mice received either AAV9-miR-298-GFP or AAV9-mock-GFP through intravenous injection. Total miRNA was collected from lumbar spinal cord and quadriceps muscle at 2,4,8 and 12 weeks after injection. qRT-PCR was performed to estimate expression level of miR-298 in (A) quadriceps muscle (n = 5) (B) lumbar spinal cord (n = 5, P <0.01). All data are reported as means ± standard error mean. The widespread transduction of the AAV vector in tissues harvested at 10 weeks after treatment was confirmed by localization of staining for GFP in the (C) quadriceps muscle (original magnification, 10X. Scale bar = 100 µm) and (D) motor neurons in lumbar spinal cord (original magnification, 40X. Scale bar = 10 µm. GFP (green), ChAT (red) and DAPI (blue). The figure has been modified from doi: 10.1038/mt.2016.13.14 Please click here to view a larger version of this figure.
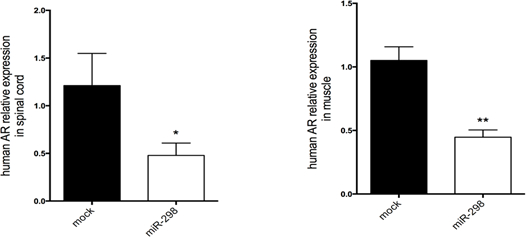
Figure 3: MiRNA-298 over-expression downregulates mutant AR in spinal cord and quadriceps muscle in mice. qRT-PCR was performed to estimate expression levels of AR mRNA in (A) lumbar spinal cord and (B) quadriceps muscle treated with AAV9-miR-298-GFP or AAV9-mock-GFP. Transcript levels were normalized to snoRNA202 (n = 5 per treatment). *P <0.05, **P <0.01. All data are reported as means ± standard errors. The figure has been modified from doi: 10.1038/mt.2016.13.14
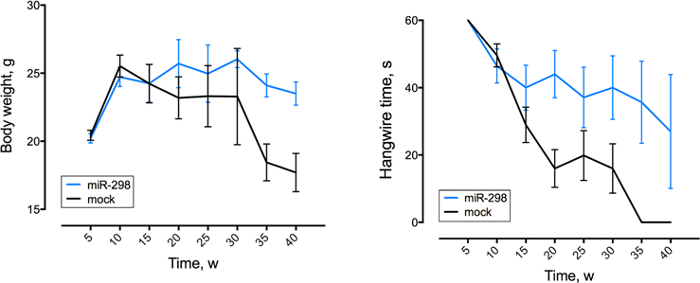
Figure 4: MiR-298 over-expression improves motor function and reduces weight loss. Behavioral assessment was performed once a week (w), between week 5 to 40. Body weight (left) and hanging wire (right) performance of mice (n = 15 per group). All data are reported as means ± standard errors. The figure has been modified from doi: 10.1038/mt.2016.13.14
