High calcium induced terminal differentiation of neonatal and adult KCs. The primary mouse epidermal KCs plated and maintained at 0.06 mM CaCl2 grew as a monolayer, and individual cells had a polygonal shape with distinct intercellular space, showing a cobblestone appearance when confluent (Figure 1A and Figure 2A). Elevating the CaCl2 to 0.2 mM induced a rapid morphology change of the cells. Within 8 h after the high calcium switch, the cells became flattened and the distinct intercellular space became less apparent, and by 24 h the cell-cell adhesion with tight junctions became obvious (Figure 1A and Figure 2B). The formation of the cornified cell envelope and vertical cell stratification started around 48-72 h after the high calcium switch (Figure 2B). To analyze the actin remodeling and cell-cell tight junction formation during the high calcium switch, KCs treated with 0.2 mM CaCl2 were stained with phalloidin (Figure 1B) to measure the actin remodeling during KC differentiation13. The formation of the actin fiber-rich filopodial projections between the adjacent cells were detected as early as 3 h post calcium switch, and the actin fiber remodeling further progressed between 6-24 h, and by 48 h the cell stratification became prominent (Figure 1B).
Susceptibility of mouse KCs to UVB-triggered cell death and TNFα release. The cultured adult mouse KCs were exposed to 25 mJ/cm2 UVB irradiation, and the cell viability and TNFα released in the culture medium were analyzed. As shown by the phase contrast images (Figure 3A), as well as the cell viability assay (Figure 3B), the UVB triggered a time dependent death of the KCs. By 24 h, the majority of the cells were rounded and were detached from the dish (Figure 3A). As quantified by the cell viability assay, ~90% of cells were dead within 24 h post UVB treatment (Figure 3B; p <0.001 as calculated by one-way ANOVA multiple comparison test). Finally, we showed that TNFα, an important cytokine that is induced by UVB and drives KC apoptosis following UVB irradiation14, was abundantly secreted (~250 pg/mL within 24 h post UVB-irradiation; p <0.001) in the culture medium of UVB-treated KCs in a time dependent manner (Figure 3C).
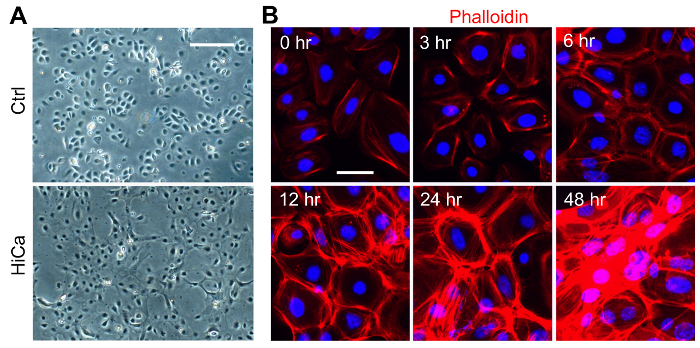
Figure 1: Primary Culture of Neonatal Mouse KCs, and High Calcium Induced Terminal Differentiation and Tight Cell-cell Junction Formation. (A). Primary neonatal mouse KCs were treated with high calcium (0.2 mM CaCl2), and phase contrast images at 4X magnification were taken at 0 hour (ctrl) and at 8 h after treatment. Scale bar = 100 µm. (B). Neonatal KCs were differentiated in the presence of 0.2 mM CaCl2, and the cells were stained with phalloidin (red) to visualize the formation of actin fiber-rich filopodial projections between the adjacent cells during differentiation. Nuclei were counterstained with DAPI, and actin fibers were stained with rhodamine phalloidin. Scale bar = 25 µm. Images were taken at 20X magnification using a fluorescence microscope set to the DAPI channel (for nuclei staining) and RFP channel (for actin staining). Please click here to view a larger version of this figure.
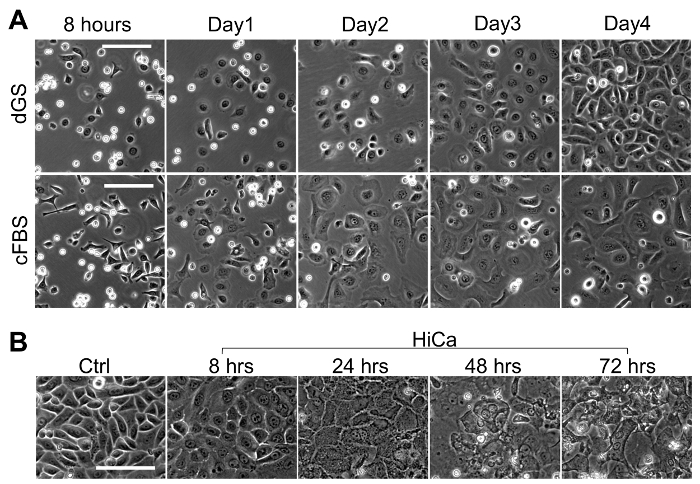
Figure 2: Primary Culture and High Calcium Induced Terminal Differentiation of Adult mouse KCs. (A). Phase contrast images at 10x magnification of primary adult mouse KCs at 8 h, 1 day, 2 days and 3 days after the initial plating. The upper panel represents cells grown in dGS, and the lower panel represents cells grown in 10% chelexed FBS (cFBS) with 8 ng/mL recombinant mouse EGF. Scale bar = 100 µm. (B). Confluent adult mouse KCs were differentiated in the presence of 0.2 mM CaCl2, and phase contrast images at 10X magnification were taken at 0 (ctrl), 8, 24, 48, and 72 h post high calcium switch. Scale bar = 100 µm. Please click here to view a larger version of this figure.
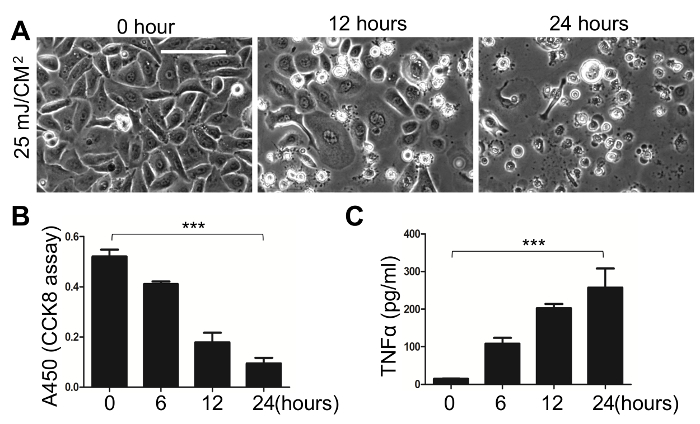
Figure 3: Primary Culture of Adult mouse KCs, and Susceptibility of Adult KCs to UVB-induced Cell Death and Release of TNFα. Primary adult mouse KCs were cultured to confluency, then exposed to 25 mJ/cm2 UVB irradiation. (A) Phase contrast images at 10X magnification taken at 12 and 24 hours post-UVB exposure showed a time dependent UVB-induced cell death. Scale bar = 100 µm. (B) The cell viability was quantified by the CCK-8 cell viability assay at 6, 8, and 24 h post-UVB treatment. (C) UVB-induced release of TNFα to the media at indicated time points was measured by ELISA. All error bars indicate mean ± SEM. The p-values were calculated by one-way ANOVA multiple comparison test (***, p <0.001) Please click here to view a larger version of this figure.
