Multiple aspects of neuroplasticity and neuron-environment interaction can be investigated using DRG and a single dissociated cell culture model. We began the studies by isolating a DRG explant and DRG-derived dissociated cells as schematically represented in Figure 1. Both tissue and single cells models can be analyzed by using a variety of molecular techniques such as immunofluorescence, Western blot, genomic assays, and other analytical techniques depending on the nature of experimental design and aims. First, we used our DRG explant model to investigate the effect of biochemical cues on axonal growth (Figure 2). In double or triple immunofluorescence, the differential expression of peripherin (Figure 2A, B; red) and β-tubulin (Figure 2A, B; green) were confirmed both within the neuronal cell bodies and the peripheral neurite sprouts. Cell nuclei were identified with Hoechst stain (Figure 2A; blue). The expression of the above-mentioned markers was further analyzed in the entire DRG explant after fixation in 4% paraformaldehyde and microtome-sectioning (Figure 3A). Our results confirmed that peripherin and β-tubulin are selectively expressed in two subpopulations of ganglionic neurons with peripherin being expressed in the "small, light" and β-tubulin in the "large, dark" subpopulations of neurons, respectively. The neurites immunolabeled with either marker emerging from the sensory neurons were found coursing through the entire ganglion before exiting into the periphery. Hoechst stain mostly identified the position of the glial satellite nuclei within the ganglion. As indicated in Figure 3B, the neuron-satellite cell interaction can be better visualized after the full cell dissociation technique. Blue-stained satellite cell nuclei were seen surrounding a β-tubulin-positive sensory neuronal soma and its extensive neurites.
We further utilized the DRG-derived dissociated cells to investigate HSV-1 infection and associated inflammatory response within the sensory neurons (Figure 4). After only 1 h of post infection (p.i.), HSV-1 entry was detected by using anti-HSV-1 gD antibody both in glial cells (Figure 4A) and DRG neurons (Figure 4B). In addition, HS staining was further carried out since HS provides a docking sites for virus attachment or binding to host cell. HS has been known as a critical component of the multi-functional extracellular matrix (ECM) and extensively documented to play a major role in cell adhesion, cell-to-cell signaling, and ECM remodeling during wound healing, embryonic development, cancer invasion, fibrosis, and in neuroinflammation35,36. Interestingly, we observed HS staining in the ECM suggesting its potential role in the microenvironment that allows for axonal growth (Figure 4C). Finally, for β-tubulin staining we used as a positive marker for the neurites to confirm the integrity of the neurons.
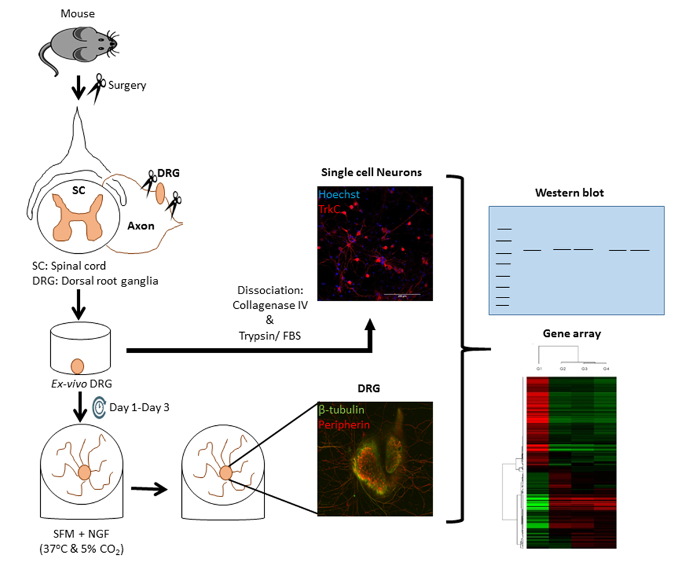
Figure 1. Schematic representation of the experimental DRG and single cell culture models. DRG were isolated from Adult NIH/SWISS mice, cleaned from excessive capsular connective tissue, and either explanted ex vivo as a whole explant or dissociated into primary co-cultures of sensory neurons and satellite cells. Both models can be used for further analyses with multiple analytical techniques. Please click here to view a larger version of this figure.
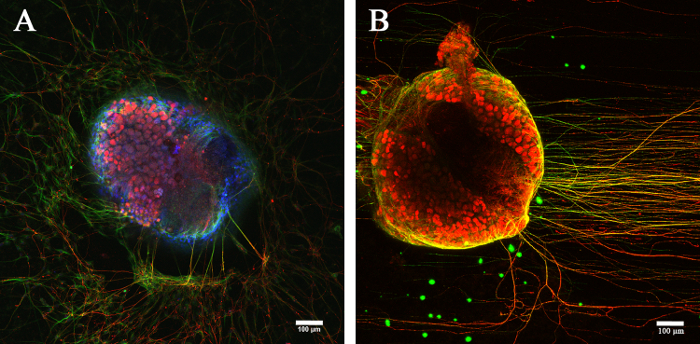
Figure 2. Development of DRG organotypic culture as a model system to investigate axonal growth. DRG grown for 6 days ex vivo were used to investigate axonal sprouting from the explant. We show by immunofluorescence how different biochemical conditions can affect the distribution of fiber sprouts: growing randomly and disorganized ( A) compared to a more linear and organized fashion ( B). The explants are labeled with anti-β-Tubulin (green) and anti-peripherin (red). Cell nuclei are stained with Hoechst stain (blue). Scale bars = 100 µm. Please click here to view a larger version of this figure.
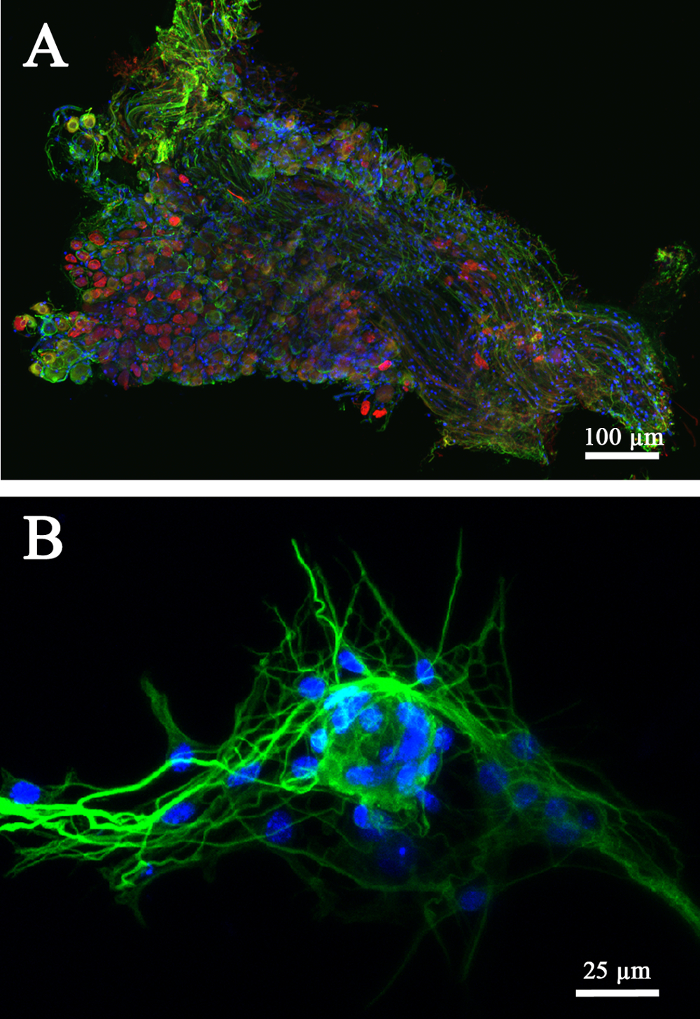
Figure 3. Molecular characterization of the isolated neuronal culture model. Sections of the whole DRG were immunolabeled with antibodies against peripherin and β-tubulin showing selective expression of the two markers in two subpopulations of ganglionic neurons. Peripherin (red) is expressed in the "small, light" neurons and β-tubulin (green) is expressed in the "large, dark" neurons. Hoechst stain mostly identified the position of the glial satellite nuclei within the ganglion ( A). The neuron-satellite cell interaction can be better visualized after full cell dissociation technique ( B). Here we show a β-tubulin-positive sensory neuronal body and its neurites surrounded by satellite glial cells (Hoechst: nuclei, blue). Scale bars, A = 100 µm; B = 25 µm. Please click here to view a larger version of this figure.
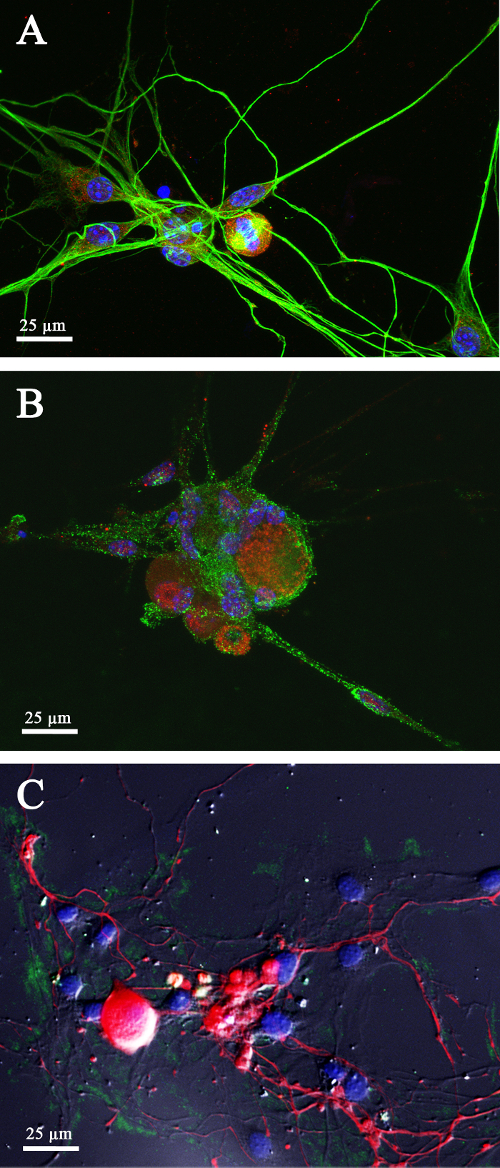
Figure 4. Investigation of HSV-1 entry model in DRG-derived dissociated cells. HSV-1 infection of dissociated satellite cells depicted along β-tubulin-positive neurites (green) is revealed using an anti-viral gD antibody (red). Satellite glial cells nuclei are stained blue with Hoechst ( A). The same antibody reveals viral entry in dissociated DRG neurons ( B). Cells are co-labeled with an antibody against heparan sulfate (HS, green) expressed on the cell membrane. HS is also a component of the extracellular matrix (green) and plays an important role for axonal growth ( C). Neurons are labeled with peripherin (red) while Hoechst blue is used to identify satellite cell's nuclei. Scale bars = 25 µm. Please click here to view a larger version of this figure.
