Gelatin-filling of the jejunum
A typical procedure for filling mouse jejunum with gelatin is shown (Figure 1A), as are the benefits of gelatin-filling (Figure 1B). In brief, the beveled tip of an 18-gauge straight needle was removed to protect against piercing the gut wall (Figure 1A1). Gelatin filling was achieved using 10% buffered neutral formalin solution injected from one end of a clipped jejunum section to flush the intestine and to fix the gut lumen surface (Figure 1A2) prior to filling with liquid gelatin (Figure 1A3). The resulting sausage-like pieces (Figure 1A4) were embedded, frozen, and transversely sectioned into 30 µm-thick slices in a cryostat. In the absence of gelatin filling, jejunal sections tend to kink, allowing the villi to swing backward (Figure 1B, left). Gelatin filling preserves the round disc-shape of the sections and maintains upright positioning of the villi (Figure 1B, right). The images clearly indicate the benefit afforded by gelatin filling in preserving the morphology of free-floating jejunum sections.
Successful imaging of intestinal tuft cells
pY1798 antibodies can be applied for variable immunostaining techniques, including dot blotting, western blotting, paraffin section staining, thaw-mounted cryosection staining, or free-floating cryosection staining1,9. In this protocol, we focused on free-floating cryosections for fluorescence staining of TCs in mouse jejunum using pY1798 antibodies, phalloidin, and DAPI staining to demonstrate the structural characteristics of TCs1. In general, TCs are scattered at a rate of approximately one TC/100 epithelial cells from villus tip to crypt1. The representative results show that pY1798 reproducibly delineates entire TCs, including the membrane, cytoplasm of the spool-shaped soma, and the strongly stained lumenal tip, where robust signal condensation corresponds to the protruding 'tuft' of a TC1 (Figure 2A-2B). Meanwhile, phalloidin is a phallotoxin, a family of poisonous bicyclic heptapeptides from the mushroom Amanita phalloides, and has high affinity for filamentous actin (F-actin), which is present in microvilli that form the intestinal brush border11. Phalloidin reproducibly and prominently marks the thickened brush border that corresponds to a mass of rootlets extending from the tuft1. Thus, the consistent co-localization of pY1798 signals (spool-shaped soma, signal condensation at the lumenal tip) with the prominently thickened phalloidin-positive brush border demonstrates that this protocol successfully identifies TCs regardless of whether they are located on a villus (Figure 2A) or in a crypt (Figure 2B).
Low-temperature antigen retrieval is effective for staining free-floating sections
Heat-based antigen retrieval at 95 – 99 °C is widely used for analysis of paraffin sections and slide-mounted cryosections. However, applying this approach to free-floating sections without causing damage is difficult because such sections are generally more fragile than slide-mounted sections that are supported by the slide12. Since low-temperature antigen retrieval (50 °C for 3 h) using a water bath did not affect the morphology of the free-floating jejunum sections in preliminary experiments, we evaluated the objective effectiveness of antigen retrieval in a blind test, which statistically confirmed the effectiveness of low-temperature antigen retrieval (Figure 3).
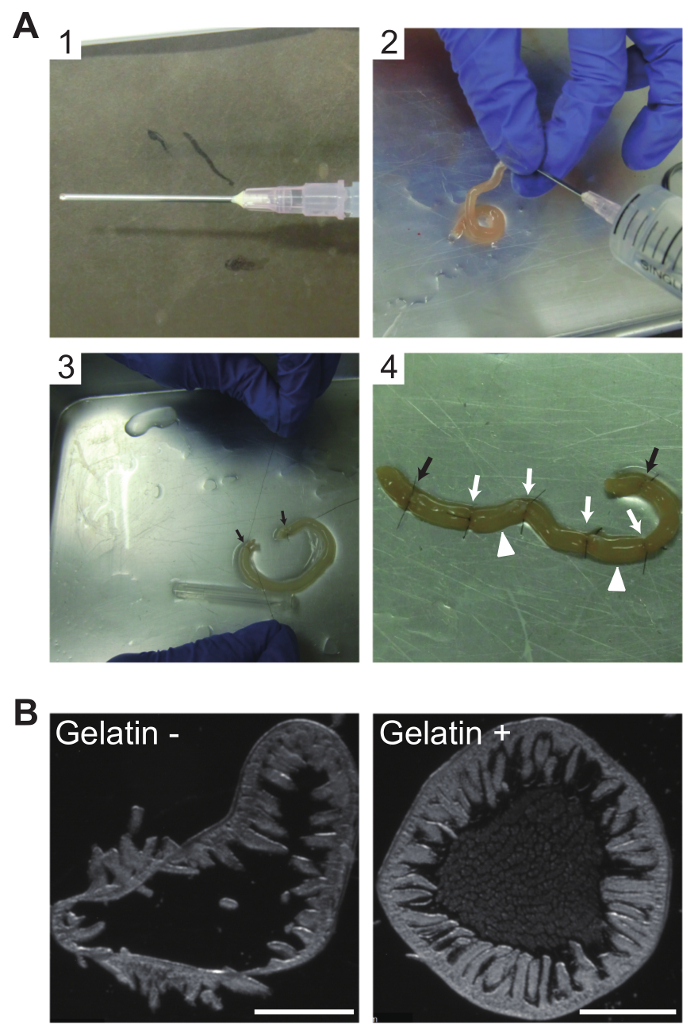
Figure 1: Gelatin filling of jejunum sections for morphological preservation of cryosections
(A) Photographs of procedures for intraluminal filling of mouse jejunum with gelatin. (A1) The beveled tip of an 18-gauge straight needle was removed to avoid piercing the gut wall. (A2) Buffered neutral formalin solution (10%) was injected in one end of the clipped jejunum to flush the intestinal contents and fix the gut lumen surface. (A3) Clipped jejunum filled with liquid gelatin and both ends ligated using a 6-0 nylon suture (black arrows). (A4) Four more suture knots (white arrows) placed between the pre-existing suture knots (black arrows) yielded three shorter pieces. The tissue will then be separated into three sausage-like pieces at the two indicated positions (white arrowheads). (B) Beneficial effects of gelatin-filling on free-floating cryosection morphology. Without gelatin filling, sections tend to kink, allowing the villi to easily swing backward (left); with gelatin filling, a 30 µm-thick section maintains a disc shape and the upright position of the villi is preserved (right). Images were photographed using Nomarski differential interference contrast. Scale bars, 1 mm. Please click here to view a larger version of this figure.
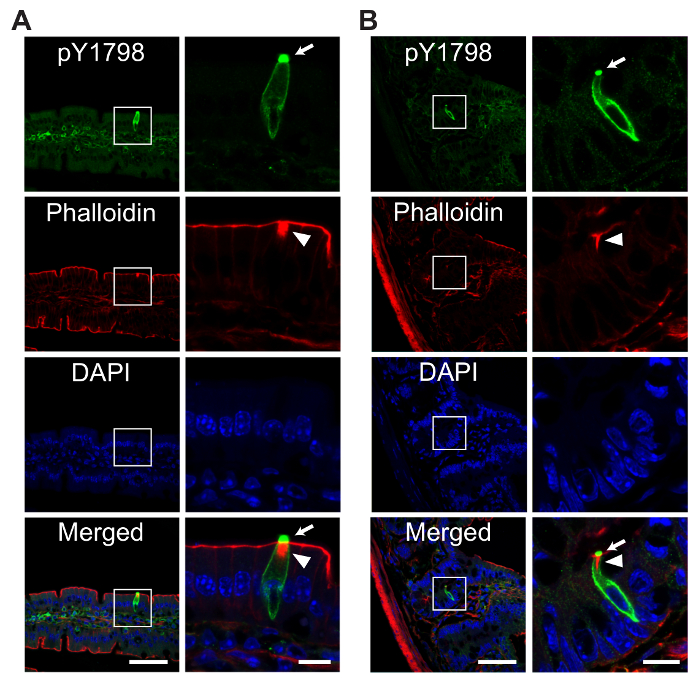
Figure 2: Representative fluorescence images of mouse intestinal tuft cells
Confocal fluorescence images of TCs on a villus (A) or in a crypt (B), in free-floating mouse jejunum sections stained with site-specific and phosphorylation-status-specific antibodies against girdin phosphorylated at tyrosine 1798 (pY1798, green, optimal excitation/emission wavelengths 490/525 nm), phalloidin (red, 590/617 nm), 4,6-diamidino-2-phenylindole (DAPI, blue, 358/461 nm). The area enclosed by white boxes in the low magnification images (scale bars, 50 µm) is expanded on the right (scale bars, 10 µm). pY1798 antibodies reproducibly stain TCs, regardless of location (on a villus (A), or in a crypt (B)), with staining present at the lumenal tip (arrows), membrane, and cytoplasm of the spool-shaped TC soma. A prominently thickened brush border in phalloidin staining (arrowheads) is another distinctive sign of TCs. Please click here to view a larger version of this figure.
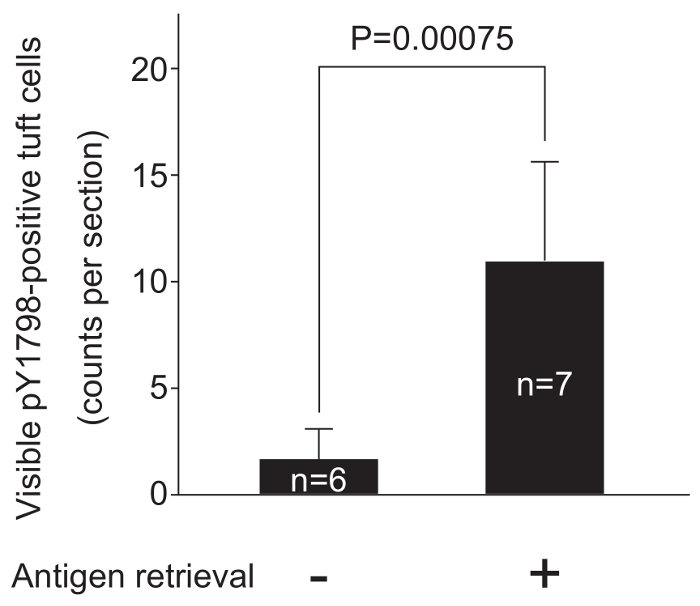
Figure 3: Low-temperature antigen retrieval enhances pY1798 immunofluorescence
Two groups of experimental procedures were compared to confirm the effectiveness of low-temperature antigen retrieval. Free-floating jejunum sections (30 µm thick, n = 13) from a single frozen block were divided into two groups, and sections from each group were stained following the complete protocol or the same protocol lacking low-temperature antigen retrieval (step 4.2.2). After staining, sections were mounted on individual slides labelled with group numbers, and the labelling on each slide was covered with masking tape. All slides were shuffled, labelled with new numbers on the tape, and observed under fluorescence microscopy through an FITC filter. Total counts of visible pY1798-positive TCs were averaged in each group, and were displayed in a bar graph (mean ± standard deviation). A two-tailed t-test was performed to compare the average counts between the two groups. P-values below 0.05 were considered statistically significant. Low-temperature antigen retrieval significantly improved the effectiveness of pY1798-immunofluorescence. Please click here to view a larger version of this figure.
