Here, we present a summary of representative results obtained from the quantification of the repertoire of natural anti-glycan antibodies in a population of 20 BALB/c mice. The glycochips used in this study contained 419 different glycan structures. Most glycans were synthesized as -CH2CH2CH2NH2 spacer-armed O-glycosides, in several cases as -CH2CH2NH2 or -NHCOCH2NH2 glycosides. All glycan structures were characterized by high resolution (700- or 800 MHz) NMR spectroscopy, purified and tested by HPLC, indicating their >95% purity. We have simultaneously determined IgM + IgG anti-glycan antibodies due to a restriction in the amount of mouse serum. In the PGA, we considered values above 4,000 RFU as a positive signal of antibody binding (this value is ~10% of the top glycans RFU). The results presented in this work follow most of the guidelines for reporting glycan microarray-based data39. Only 17% of carbohydrate structures demonstrated ≥4,000 RFU in the PGA (Figure 2, in red). Most of the glycan structures exposed in the glycochips were not recognized by the repertoire of circulating anti-glycan antibodies of BALB/c mice (Figure 2, in blue and white)28. The conserved pattern of natural anti-carbohydrate antibodies of BALB/c included 12 different glycan specificities, with very high median signal intensities of antibodies binding (≥10,000 RFU Table 1)28.
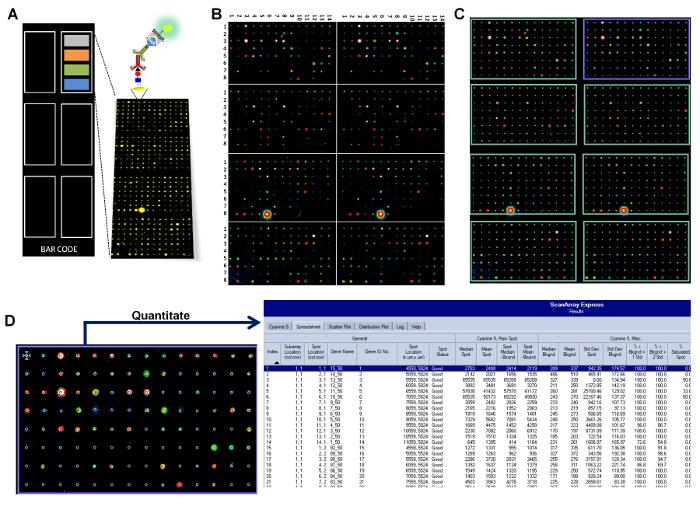
Figure 1: Schematic representation (not at scale) of the glycan array configuration, printing, and analysis. (A) Printed microchips are developed with a library of 419 different glycan structures, followed by the detection with an appropriate fluorescently labeled secondary antibody. Each slide contains 4 different blocks of sub-arrays (in colors), repeated 6 times. Every single sub-array is formed by 112 different glycan spots (8 rows × 14 columns), including controls. (B) A representative example of the images obtained from microchip scanning using a fluorescence scanner (third part of the image). (C) The process of aligning the "grid" to spots in every single sub-array (template adjustment during quantification). (D) The fluorescence is detected for each spot and results are transferred into a common spreadsheet file. Please click here to view a larger version of this figure.
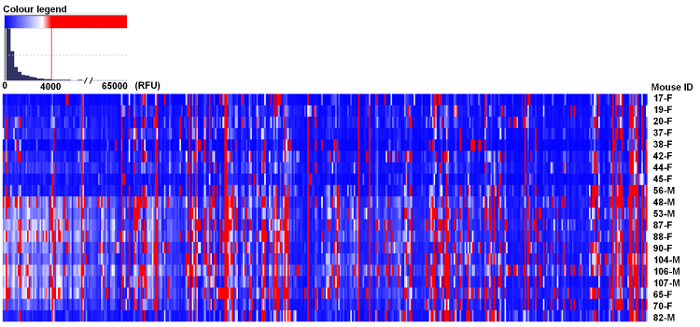
Figure 2: Repertoire of natural circulating anti-carbohydrate antibodies of BALB/c mice. Mouse serum samples (1:20) were incubated with the glycochips and scanned using a ScanArray reader. Data were analyzed with a microarray analysis system and results were expressed in relative fluorescence units (RFU) as the median ± median absolute deviation (MAD). Blue and white colors represent binding signals lower than 4,000 RFU (background); red color represents signals ≥4,000 RFU (positive binding). F: female; M: male (n = 20). This figure has been reproduced from Bello-Gil, D. et al.28. Please click here to view a larger version of this figure.
| Glycan ID (#) |
Structure | Common name | Median and MAD as RFU | Number of mice showing RFU ≥4000 (%) | |||
| 60 | 6-O-Su-Galβ-spb | 61113 | 1156 | 100 | |||
| 271 | Galβ1-6Galβ1-4Glcβ-sp | 53622 | 1934 | 100 | |||
| 802 | Galβ1-3GalNAc(furc)β-sp | 51348 | 2324 | 100 | |||
| 176 | 3-O-Su-Galβ1-4(6-O-Su)Glcβ-sp | 43008 | 9342 | 100 | |||
| 166 | GlcAβ1-6Galβ-sp | 39105 | 2993 | 85 | |||
| 150 | 3-O-Su-Galβ1-3GalNAcα-sp | 37943 | 3232 | 100 | |||
| 437 | GalNAcα1-3(Fucα1-2)Galβ1-3GalNAcβ-sp | A(type 4) | 33886 | 3193 | 90 | ||
| 125 | 6-Bn-Galβ1-4GlcNAcβ-sp | 32674 | 5389 | 95 | |||
| 154 | 3-O-Su-Galβ1-3GlcNAcβ-sp | 32651 | 3954 | 100 | |||
| 177 | 3-O-Su-Galβ1-4(6-O-Su)GlcNAcβ-sp | 32496 | 7215 | 100 | |||
| 287 | 3-O-Su-Galβ1-3(Fucα1-4)GlcNAcβ-sp | SuLea | 20063 | 4962 | 95 | ||
| 234 | Galβ1-4(Fucα1-3)GlcNAcβ-sp | Lex | 13573 | 2635 | 80 | ||
Table 1: Top rank glycan structures recognized by natural antibodies of BALB/c mice. Glycans with binding signals above 4,000 RFU in at least 80% of examined mice (n = 20). bsp means aminoethyl, aminopropyl or glycyl spacer. cfuranose; all other monosaccharides are in a pyranose form; Fuc residue has L-configuration, all other monosaccharides – D-configuration. This table has been modified from Bello-Gil, D. et al.28.
Supplementary Table 1: List of glycans, their binding to natural circulating antibodies (IgM + IgG) of BALB/c mice (n = 20), expressed in relative fluorescence units (RFU) as median ± MAD, and the number of animals exceeding cut off (≥4000 RFU). This table has been reproduced from Bello-Gil, D. et al.28. Please click here to download this table
