Shigella invasion is associated with atypical long-lasting local Ca2+ responses:
Following the protocol mentioned above, Fluo-4-loaded HeLa cells were challenged with WT Shigella and stream acquisitions were performed to analyze Ca2+ signals. A representative experiment is shown in Figure 1, with time-lapse images series of the fluorescence intensity of the Fluo-4 probe averaged in a region of interest for a single cell, and the corresponding phase contrast image (Figure 1A, left panel). The Shigella invasion site is characterized by membrane ruffles detected in the phase contrast image (Figure 1A, arrow). Atypical local increases in free cytosolic Ca2+ are observed at the Shigella invasion site with a varying amplitude and durations ranging from 2.5 – 5 s (Figures 1A and 1B, arrows), followed by global increases in the infected cell (Figure 1B, arrowheads).
The type III effector IpgD regulates the transition from local to global Ca2+ responses induced by Shigella in HeLa cells:
The analysis of Ca2+ signals induced by wild-type Shigella and an isogenic mutant strain deficient for the type III effector IpgD, a phosphatidyl 4, 5 bisphosphate phosphatase, indicated that this latter strain induced more global and less atypical local responses with long durations (RATPs) with 11.3 ± 4.4 (SEM)% and 40.3 ± 7.5 (SEM)% of cells responsive with global responses in the case of a WT and ipgD mutant, respectively (Figure 1D)15. This ipgD mutant obtains local responses at a similar frequency as the WT strain.
Shigella inhibit InsP3-dependent global Ca2+ increases in HeLa cells during prolonged infection kinetics:
The imaging of global Ca2+ responses over extended infection kinetics pointed to a decrease in the frequency of responses 30 min following the challenge with wild-type Shigella (Figure 2A). A similar decrease in the frequency of Ca2+ responses was observed in TC-7 cells infected for 30 min with wild-type Shigella15. The quantification of dose-dependent cell responses to the Ca2+ agonist histamine illustrates the inhibition of an InsP3-dependent Ca2+ release at late stages of a Shigella infection. WT Shigella leads to atypical isolated Ca2+ responses during the first 30 min of the infection and after further incubation, a drastic decrease in the amplitude and frequency of Ca2+ responses was observed (Figure 2B).
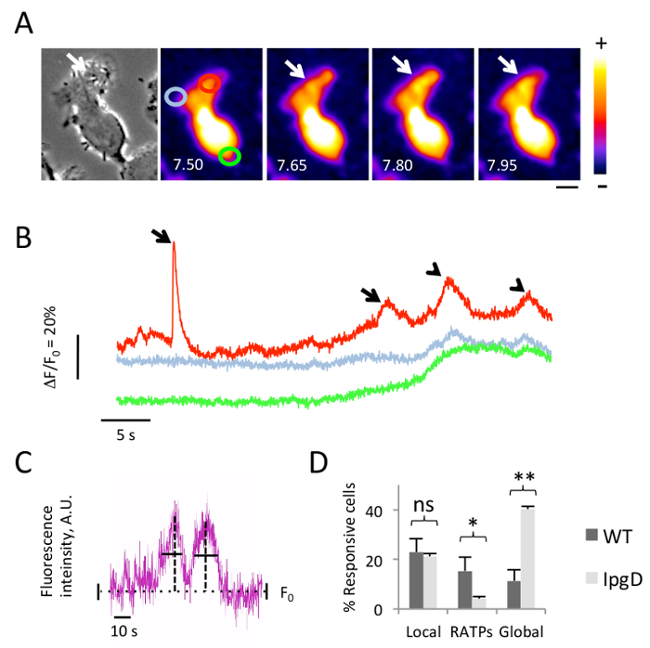
Figure 1. Local and global Ca2+ responses during Shigella invasion. HeLa cells were loaded with Fluo-4-AM and challenged with WT Shigella. (A) The left panel shows phase-contrast images. The right panel shows time series of Fluo-4 fluorescence average intensity with the color code depicted on the right. The time from the start of acquisition is indicated in seconds, 10 min after the bacterial challenge. The arrow indicates the invasion foci. The circles depicted correspond to the regions analyzed in (B) where the traces with the corresponding color represent the variations in Fluo-4 average fluorescence intensity over the baseline. The arrows indicate local responses. The arrowheads indicate global responses. (C) This is a scheme depicting the determination of the duration of the Ca2+ responses. The horizontal dotted line indicates the basal Ca2+ levels (F0); the vertical bars indicate background variations. The vertical dotted line indicates the maximal amplitude of the response; the horizontal bars indicate the duration of the response determined at the half-maximal amplitude. (D) This is the percentage of responsive cells ± SEM showing local responses and global Ca2+ responses induced 5 min post-infection with the WT Shigella (dark gray bars) or the ipgD mutant (light gray bars). RATPs are local Ca2+ responses that last for more than 5 s. N = 4; >60 cells for each time. Wilcoxon test, *P <0.05; **P <0.01. Please click here to view a larger version of this figure.
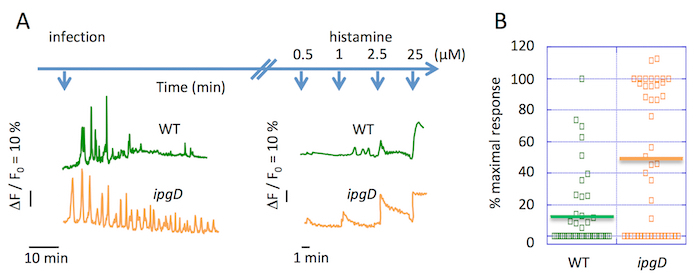
Figure 2. Inhibition of global Ca2+ responses during extended kinetics of Shigella infection. (A) The upper blue arrows represent the time scale of the bacterial and histamine challenge at the indicated concentrations. This panel shows the representative traces of single cell global Ca2+ variations following the infection by the WT Shigella (green) and ipgD mutant strain (orange). (B) This panel shows the percentage of the amplitude of Ca2+ responses relative to the maximal response, upon a stimulation at 0.5 μM histamine of the cells infected with the indicated strains for 90 min. The solid horizontal bar represents the average maximal percentage. Wilcoxon test, **P <0.01. Please click here to view a larger version of this figure.
