The size and zeta potential of PEI-SPIONs prepared with this protocol were in the range of 29 – 48 nm (polydispersity index: 0.12 – 0.23) and 30 – 48 mV, respectively. They were stable in water at 4 °C for over 12 months without obvious aggregation. To evaluate their siRNA binding ability, PEI-SPIONs were mixed with siRNA at various Fe:siRNA weight ratios. Figure 1 shows that when the Fe:siRNA weight ratio reaches 4 and above, the band of free siRNA was completely missing, implying successful siRNA binding to PEI-SPIONs. A major concern of PEI in biomedical application is its toxicity, which is a result of the strong positive charge, particularly at high molecular weights and high doses. As shown in Figure 2A, PEI-SPIONs with a zeta potential of 30.5 and 37 mV did not exhibit apparent cytotoxicity at concentrations up to 30 μg Fe/mL, which is about twofold higher than the concentration (15 μg Fe/mL) normally used for cell transfection. However, PEI-SPIONs with a zeta potential of 48 mV were toxic even at the lowest dose examined (10 μg Fe/mL). Therefore, PEI-SPIONs possess a charge-dependent toxicity. Since cationic charge is not important for the NP uptake by macrophages19, we suggest that PEI-SPIONs with an average zeta potential not higher than +37 mV are used for the siRNA transfer, although siRNA binding would decrease the charge to some extent and alleviate cytotoxicity18.
To test the potential application of PEI-SPIONs for siRNA delivery to macrophages, the in vitro transfection was performed with the murine macrophage cell line RAW 264.7. As analyzed by flow cytometry, more than 90% of the cells were transfected with fluorescently labeled PEI-SPION/siRNA complexes at 15 µg Fe/mL (Figure 2B). With respect to transfection efficiency, there was actually no difference between PEI-SPION/siRNA NPs formed at Fe:siRNA weight ratios of 4 and 8, although, under the latter condition, the NPs that were formed were smaller in size and weaker in positive charge because a lesser amount of siRNA was loaded per particle. We also assessed the effect of PEI-SPION/siRNA concentration on cellular internalization by Prussian blue staining. As shown in Figure 2C, the blue spots within the transfected cells were minimally detectable at 7.5 µg Fe/mL, but clearly visible at 15 µg Fe/mL. Interestingly, increasing the PEI-SPION/siRNA concentration to 32 µg Fe/mL did not increase the staining intensities, probably because the PEI-SPION/siRNA uptake was saturated at concentrations around 15 µg Fe/mL. In addition, the ability of PEI-SPIONs for mediating the siRNA transfer was corroborated in primary macrophages18, and the method presented here had a high siRNA transfection efficiency in rat peritoneal macrophages, equivalent to that in RAW264.7 cells. The peritoneal macrophages transfected with PEI-SPIONs harboring specific siRNA showed a significant decrease in the target mRNA level as compared with nonspecific siRNA (Figure 2D), implying that siRNA could escape from endocytosis vesicles into the cytoplasm and reach the RNAi machinery.
We previously also investigated the in vivo cellular uptake of systemically administered PEI-SPION/siRNA complexes in rats with adjuvant arthritis18. We analyzed PEI-SPION/siRNA transfection efficiency in phagocytic macrophages and nonphagocytic T lymphocytes. As shown in Figure 3, CD11b+ cells took up PEI-SPION/siRNA complexes more efficiently than CD3+ cells at any time point in all of the organs examined, indicating that PEI-SPION/siRNA NPs preferentially target macrophages18. Notably, a high-level accumulation of the NPs was observed in inflamed joints18, suggesting that the PEI-SPION can be an attractive platform for the systemic delivery of siRNA therapeutics in rheumatoid arthritis whose pathogenesis is linked to macrophage dysfunction and for which local siRNA administration is not a favorite choice due to the involvement of multiple organs of the disease.
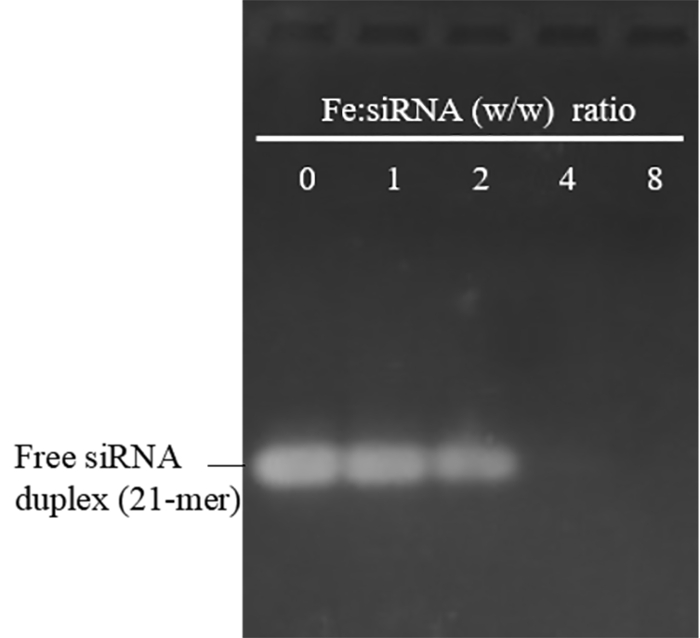
Figure 1: Agarose gel electrophoresis of PEI-SPION/siRNA complexes formed at various Fe:siRNA (w/w) ratios. A Fe:siRNA ratio of 0 represents free siRNA duplexes without PEI-SPIONS. The average size and zeta potential of the free PEI-SPIONs used here were 30 nm and 45 mV. siRNA could completely bind to PEI-SPIONs when the Fe:siRNA ratio reaches 4 and above, consistent with the previous results using PEI-SPIONs with an average size of 48 nm and a zeta potential of 30.5 mV18. The absence of retarded bands (PEI-SPION/siRNA complexes) may reflect inaccessibility of siRNA to EB during staining, an indication of strong siRNA binding, and/or the condensing ability of PEI-SPIONs. Please click here to view a larger version of this figure.
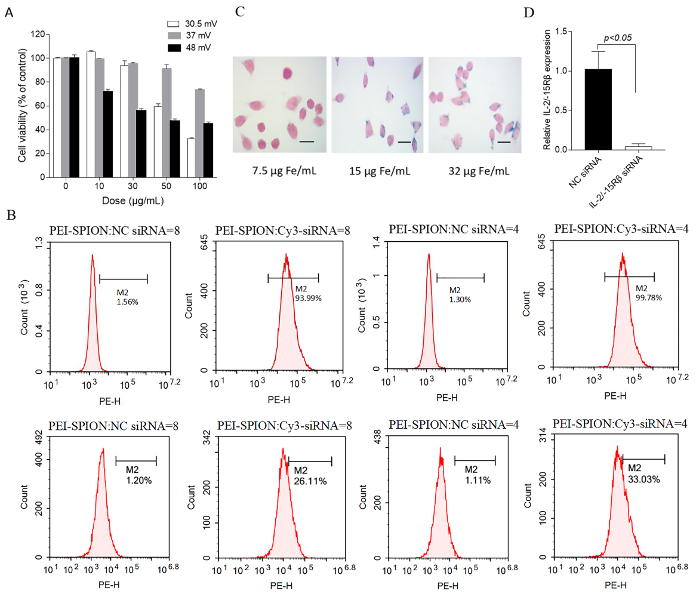
Figure 2: Biological characterization of PEI-SPION and PEI-SPION/siRNA NPs. (A) This panel shows a cell viability assay. RAW 264.7 cells were treated for 16 h with the indicated doses of PEI-SPIONs bearing different zeta potential and, then, an MTS assay was performed. The cell viability was normalized against the control (no particle exposure). The data are the mean ± SD of duplicate wells. (B) This panel shows a flow cytometric analysis of the PEI-SPION/Cy3-siRNA uptake by RAW 264.7 cells. The cells were incubated for 24 h with 15 µg Fe/mL (upper panel) or 5 μg Fe/mL (lower panel) of PEI-SPIONs complexed with Cy3-labeled siRNA at a Fe:siRNA (w/w) ratio of 4 and 8, respectively. Nonspecific (NC) siRNA represents non–fluorescent siRNA. M2: gated region; Pe-H: Cy3 fluorescence intensity. The siRNA transfection efficiency of PEI-SPIONs used here (37.8 nm, 48 mV) was similar to a previous study using PEI-SPIONs with an average size of 48 nm and a zeta potential of 30.5 mV18. (C) This panel shows an analysis of PEI-SPION/siRNA uptake by visualizing cellular iron deposits. RAW 264.7 cells were incubated with 7.5, 15, and 32 μg Fe/mL PEI-SPIONs (48 nm, 30.5 mV) complexed with siRNA (Fe:siRNA = 8) and stained by Prussian blue. The scale bars are 20 μm. (D) This panel shows an in vitro validation of the silencing efficiency of siRNA delivered by PEI-SPIONs. A specific siRNA-targeting rat IL-2/-15 receptor β chain was loaded onto PEI-SPIONs (48 nm, 30.5 mV) at Fe:siRNA = 8 and, then, transfected into rat peritoneal macrophages. An NC siRNA was used as control. The gene silencing effect was assessed by quantitative PCR. The cells were incubated with the complexes at 15 μg Fe/mL. The data are the mean ± the SD of triplicate wells. Panel D has been modified from Duan et al.18 with permission from the publisher. Please click here to view a larger version of this figure.
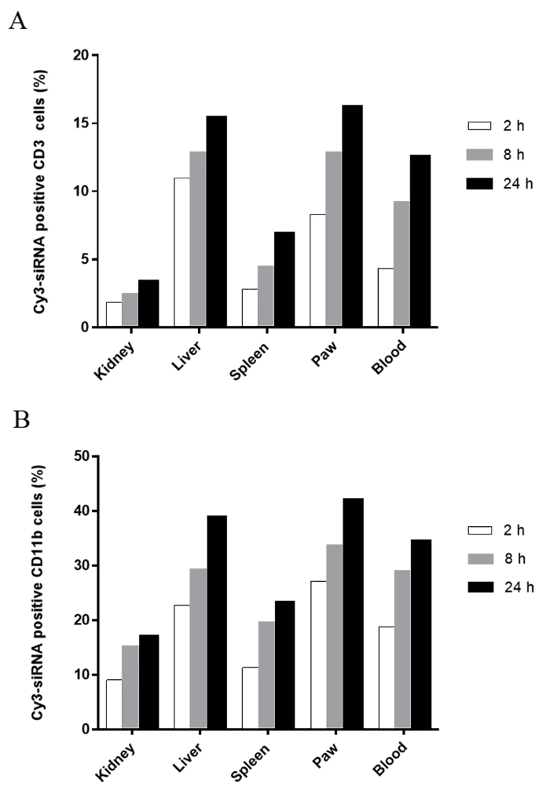
Figure 3: In vivo cellular uptake of PEI-SPION/siRNA NPs. Three arthritic rats were injected intravenously with a single dose of 0.3 mg/kg Cy3-siRNA formulated with PEI-SPION (48 nm, 30.5 mV). A rat injected with PBS was used as a control. Blood, spleen, liver, kidney, and inflamed joints were collected at 2, 8, and 24 h after the injection. The cellular uptake of PEI-SPION/Cy3-siRNA NPs was assessed by flow cytometry using (A) anti-CD3 and (B) anti-CD11b monoclonal antibodies. The percentages are of Cy3-siRNA uptake within the gated CD3+ or CD11b+ cells. The results shown here are representative of two independent experiments. This figure has been modified from Duan et al.18 with permission from the publisher. Please click here to view a larger version of this figure.
