The above protocol describes set up, inoculation, and running of a 5-stage in vitro system to study the gut microbiota of the colon. To generate the data presented below, following DNA extraction, 16S rRNA marker gene DNA sequencing of the V1V2 region was performed using the high throughput sequencing (e.g., MiSeq Illumina platform) by the Microbiome Center at the Children’s Hospital of Philadelphia27. QIIME (Quantitative Insight into Microbial Ecology) version 1.928 was used to process the sequencing data and statistical analysis was performed on the R environment for statistical computing29. Taxonomic assignments were generated using the Greengenes 16S reference database30,31. OTU relative abundance values were calculated by dividing the OTU read count by the total number of reads in the sample. SCFA levels were quantified using the protocol described.
The in vitro communities achieve a steady state equilibrium
The ability to study the gut microbiota and short chain fatty acid (SCFA) production in vitro with a multi-stage system requires that the microbial communities reach a steady state. This occurs after inoculation when the taxa have become established in their niches, and the composition of the community and their metabolites are no longer fluctuating. In a steady state, the community and its products remain the same over time. For in vitro gut microbiota studies, stability is a key requirement; without stability, it is impossible to determine whether or not observed changes are occurring due to the experimental conditions, or if they are due to variation. According to what has been proposed in the literature, a community can be considered stable when there is 80% similarity between time points32.
Using the results of 16S rRNA marker gene sequencing, Principal Coordinates Analysis (PCoA) plots based on unweighted and weighted UniFrac distances were generated for the communities that developed in each region of the system over time (Figure 2). An unweighted analysis compares the communities in terms of the presence/absence of species. The weighted analysis compares them based not only on the presence/absence of species but also considers their abundance. For Unit 1, this included both the luminal and mucosal phases, and for Unit 2 only the luminal phase. Based on this analysis, it was observed that the community changed appreciably between days 1 and 7. However, from day 11 post inoculation until the end of the experiment, the samples occupied a small region of the PCoA space. This was true for sample-sample distance scores based on OTU presence-absence (Figure 2A) or on OTU abundance (Figure 2B). Thus, it was observed that, after 11 days, the abundance and the types of bacteria were stable from one sample point to the next. This pattern was observed for both the luminal and mucosal phases of all three colon regions of Unit 1 and the luminal phase of Unit 2. These results illustrated that both the luminal and mucosal phases reach stability and that this occurred at the same time.
System stability was also probed by analyzing the production of short chain fatty acids (SCFAs) over time. The three most prominent SCFAs (propionic acid, butanoic acid, and acetic acid)38 were measured in each sample over the course of the experiment using gas chromatography GC/MS. These measurements revealed that propionic acid, butanoic acid, and acetic acid fluctuated from the start of the experiment until day 15 post inoculation (Figure 3A-C). After day 15, the amounts of these SCFAs produced in each colon region remained constant, with only minimal changes occurring until the end of the experiment (Figure 3A-C). The difference between time points was an average of 6.8% for propionic acid, 7.2% for acetic acid, and 8.02% for butanoic acid. This suggests that similar to the community composition, the metabolic properties of the community entered a steady state, as indicated by the production of stable amounts of SCFAs over time. Both Unit 1 and Unit 2 produced similar amounts of SCFAs, with no significant differences between the two (p > 0.05). This indicated that the production of SCFAs was not affected by the presence or absence of the mucosal community. It should be noted that the point of stability determined in this experiment is similar to that reported previously, in which it was stated that community stabilization in a 5-stage in vitro system occurred approximately 2 weeks post inoculation33,34,35.
The communities developed in the in vitro system are similar to the inoculum
The second required element for an in vitro gut microbiota experiment is that the community developed in the model preserves the microbial diversity of the fecal inoculum. Since the system used in this experiment provides for three distinct colon regions it cannot be expected that any one region be exactly the same as the fecal inoculum. However, it is expected that the members of each community are derived from the fecal inoculum, and all together maintain a similar level of diversity as the inoculum.
Based on the results of 16S rRNA marker gene sequencing, the average, stable community for the luminal and mucosal phase of each colon region was determined (Days 15-28 post inoculation) and compared to the community of the fecal inoculum (Figure 4A). These results demonstrate that the communities which developed in the individual colon regions of the in vitro system were similar to the fecal inoculum in composition. The two most prominent orders in the reactor, Clostridiales and Bacteroidales, matched those observed in the inoculum. However, several low-abundance bacterial orders in the colon regions were different from the inoculum, with the most prominent differences between orders Burkholderiales and Synergistales (Figure 4A).
The alpha diversity for in vitro system was compared to the inoculum, as another measure of community structure after stabilization. The Shannon index, calculated for each region over time, reached a similar level of diversity as the inoculum for all regions, except for luminal samples from the ascending region (Figure. 4B). Taken together, these results demonstrate that the in vitro culture system was able to produce a community comparable to the fecal inoculum, both in terms of composition and diversity (Figure 4A-B).
Using an in vitro system allows for the development of the region and the phase specific communities
The three colon regions represented in this system are maintained at different pH values and receive different nutrient supplies. (This was mentioned in the protocol section). Based on this, it is expected that the communities in these regions will differentiate 33. The stable (Days 15-28) luminal communities in both Unit 1 and Unit 2 were determined at the family level and plotted according to relative abundance (Figure 5). The divergence between the three colon regions in terms of the abundance of specific taxa, demonstrates that each region develops a unique community. This is also supported by the results of Figure 2 and Figure 4A. In Figure 2, the mature communities that developed in each region cluster at different locations in the PCoA chart. In Figure 4A, the communities in each colon region differed in the percentages of the dominant order members.
For this experiment, Unit 1 was provided with mucosal carriers, while Unit 2 had no mucin carriers. Based on this experimental design, the contribution of the mucosal surface could be examined. For comparison purposes, the stable (Days 15-28) luminal and mucosal communities for Unit 1, at the family level, were plotted together according to relative abundance (Figure 6). It is clear that the composition of the luminal and mucosal communities for all three colons differ in the abundance of some taxa, the most prominent being Lachnospiraceae and Bacteroidaceae. These results also demonstrate that the mucosal communities in the three regions are different from each other. For example, Clostridiaceae is enriched in the mucosal communities of the descending and transverse regions, and Veillonellaceae is higher in the mucosal phase of the ascending colon, but not in the transverse or descending colon regions. Taken together, the results presented in these figures show that there is a clear difference between the communities in each phase and each region for some taxa, illustrating that the composition between the regions is similar and that there is a difference in abundance.
Although analysis of DNA sequencing reveals the development of a region-specific community, there is no apparent difference in the production of SCFAs between regions. The average ratio of Acetic Acid: Propionic Acid: Butanoic Acid for the stable community (Day 15-28) was calculated for each region (Figure 7A). While the ratios of the different SCFA remained fairly similar, there is an increase in the total amounts of SCFAs produced between the ascending, transverse and descending regions, with the highest levels found in the descending region (Figure 7B). This was true for both Unit 1 and Unit 2.
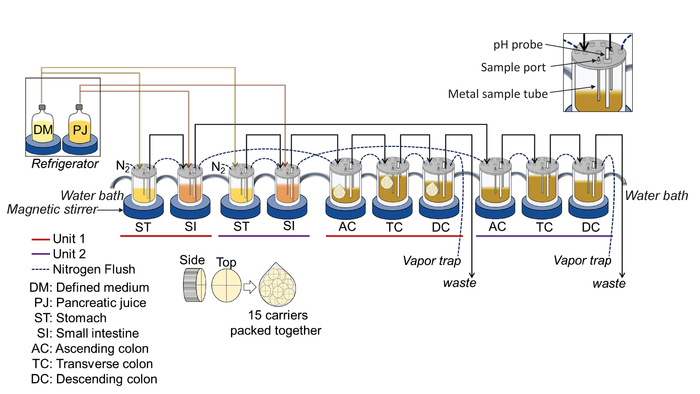
Figure 1: Illustration of the 5-stage, in vitro experimental design. The complete system consists of the following components: A circulating water bath, nitrogen flow, a set of glass bioreactors, a set of magnetic stirrer bars and magnetic stirrers, pH probes, a computer-controlled console containing 40 peristaltic pumps, a computer monitor, and a refrigerator. The main system is composed of a set of bioreactors mimicking the stomach, small intestine, and the ascending, transverse, and descending colon regions. Two complete units are set up to run in parallel, providing for an experimental and a control group. This means that 10 bioreactors, 10 pH probes, and 10 magnetic stirrers are required for this experiment. Please click here to view a larger version of this figure.
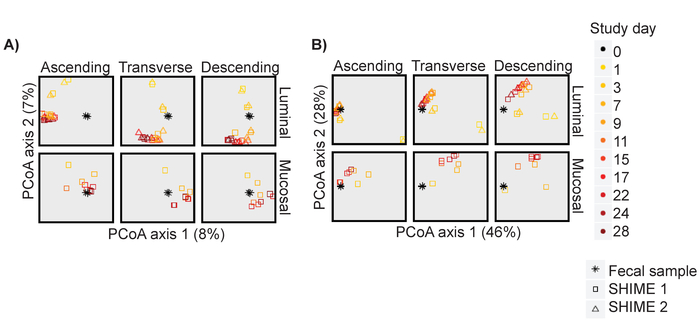
Figure 2: The in vitro community stabilizes by day 11 post inoculation. PCoA analysis based on (A) Unweighted and (B) Weighted Unifrac distances for the luminal and mucosal phase of each region over time for Unit 1 and for the luminal phase of Unit 2. The plot is faceted into the components after calculating the PCoA axes. Please click here to view a larger version of this figure.
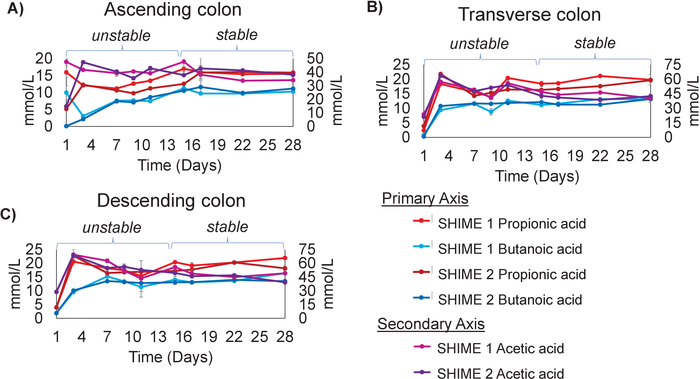
Figure 3: The production of SCFAs by the in vitro system stabilize by day 15 post inoculation. Measurements of Propionic Acid, Butanoic Acid, and Acetic Acid over time for the (A) Ascending colon (B) Transverse colon and (C) Descending colon. The experiment was performed in triplicate. The results represent an average of three independent measurements, with error bars depicting the standard deviation. Please click here to view a larger version of this figure.
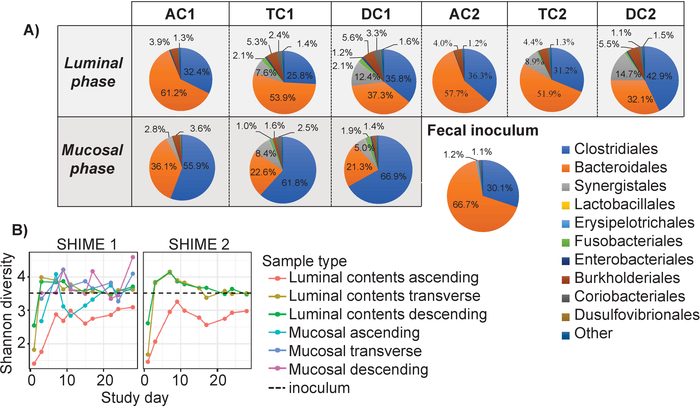
Figure 4: Comparison of the stable communities from the in vitro system to the fecal inoculum. (A) The stable communities (D15-28), at the order level were averaged and formatted in pie charts for both the mucosal and luminal phase of each colon region, and for the fecal inoculum. (B) The Shannon diversity for the mucosal and luminal phase of each colon region compared to the fecal inoculum (black dotted line). Please click here to view a larger version of this figure.
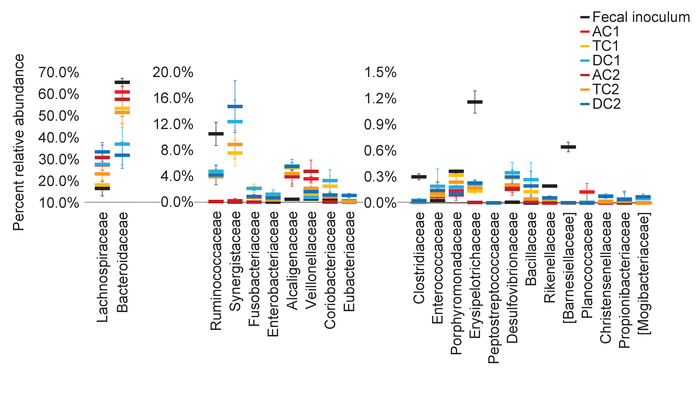
Figure 5: The luminal and mucosal phase in each colon region promote the growth of distinct communities. The average relative abundance, at the family level, for the stable communities (D15-28) for the luminal and mucosal phase, and the inoculum, were calculated and plotted together for each colon region. Error bars represent the standard deviation between timepoints. AC1 = Ascending colon Unit 1; TC1 = Transverse colon Unit 1; DC1 = Descending colon Unit 1; AC2 = Ascending colon Unit 2; TC2 = Transverse colon Unit 2; DC2 = Descending colon Unit 2. Please click here to view a larger version of this figure.
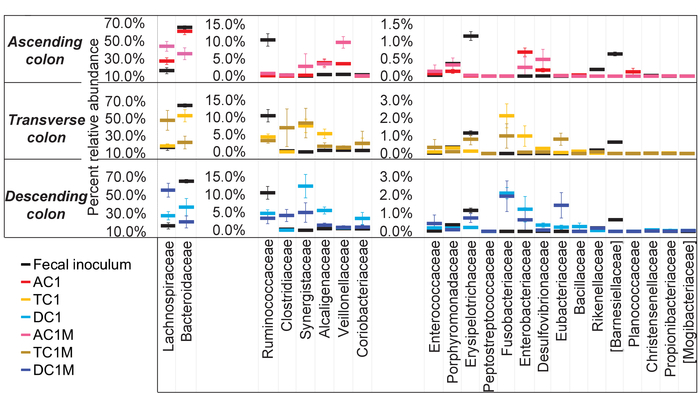
Figure 6: Each colon region of the in vitro system develops a unique community. The average relative abundance, at the family level, for the stable communities (D15-28) in each colon region, and the fecal inoculum, were calculated and plotted together. The experiment was performed in triplicate. The results represent an average of three independent measurements, with error bars depicting the standard deviation. Please click here to view a larger version of this figure.
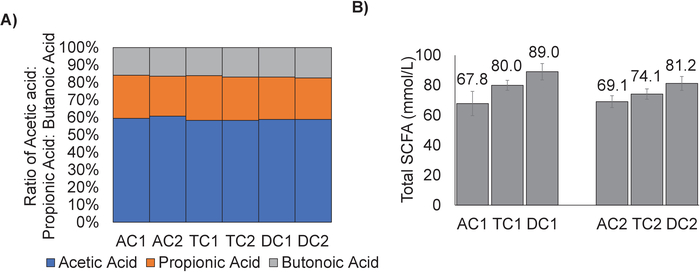
Figure 7: The ratio of Acetic Acid: Propionic Acid: Butanoic Acid is similar for each colon region. (A) The amounts of acetic acid, propionic acid, and butanoic acid for the stable communities (D15-28) of each colon region were calculated and converted to a ratio. (B) The ratios were plotted as percentages for each colon region. Error bars represent the standard deviation between timepoints. AC1= Ascending colon Unit 1; TC1 = Transverse colon Unit 1; DC1 = Descending colon Unit 1; AC2 = Ascending colon Unit 2; TC2 = Transverse colon Unit 2; DC2 = Descending colon Unit 2. Please click here to view a larger version of this figure.
