Human enteroid and colonoid cultures are grown as 3D structures, then dissociated and fragmented for plating on human collagen IV-coated cell culture inserts. The progress of monolayer formation is easily monitored on a daily basis via bright field microscopy, immunofluorescence staining (Figure 1), and by a steady increase in transepithelial electrical resistance (TER) (Figure 2), which reflects the permeability of tight junctions to ions and correlates with monolayer confluency. The TER of an empty 24-well insert is approximately 50-100 Ω·cm2, and upon reaching confluency increases to approximately 400-500 Ω·cm2. All epithelial cells in confluent monolayers are connected by junctional complexes detected by the F-actin ring at the cell perimeter (Figure 1). Monolayers in full growth factor media represent the proliferative crypt-like epithelium composed primarily of actively dividing cells that incorporate the nucleoside analog EdU (Figure 2). Withdrawal of the growth factors WNT3A and RSPO1 promotes differentiation, leading to villus-like cultures that lack proliferating cells. Differentiated monolayers contain enterocytes (enteroids) or colonocytes (colonoids) and develop specialized intestinal epithelial cells as has been shown in 3D cultures,18 including goblet and enteroendocrine cells.15 Differentiated monolayers also demonstrate a significant increase in TER (> 1000 Ω·cm2) (Figure 2), indicating mature tight junctions.
In normal human physiology, the intestinal epithelial layer separates nutrients and the microbe-enriched luminal space from the sterile serosal environment. The epithelium tightly regulates the cross-talk between these two compartments. Enteroid and colonoid monolayers preserve this important compartmentalization property as shown by the proteomic analysis in Table 1. There are substantial differences between the protein composition in apical and basolateral conditioned media collected from differentiated enteroid monolayers. Access to both the apical and basolateral sides allows for collection of both supernatants in a time dependent manner for measurement of differential secretion of other molecules such as cytokines and chemokines.15,17
Monolayers are convenient and highly reproducible models to detect the changes in protein expression using both immunoblotting and immunostaining. Thus, changes in mucin 2 (MUC2) expression by shRNA knockdown (KD) can be detected using both techniques (Figure 3). The shRNA transduction was performed on 3D colonoids and maintained in antibiotic selection media. After verifying the KD, colonoids can be continuously maintained as 3D cultures and/or plated as monolayers for experimental purposes. Moreover, MUC2 KD does not affect the efficiency of monolayer formation compared to the wild type parental colonoid line. Collection of monolayers for immunoblotting is simple and similar to the protocols described for human epithelial adenocarcinoma-derived cell lines. Typically, approximately 50 µg or more of total protein can be extracted from a single 0.33 cm2 insert-grown monolayer. As shown in Figure 3, MUC2 is barely detectable in undifferentiated (UD) WT colonoid monolayers but is highly expressed in differentiated (DF) WT monolayers. MUC2 is below the level of detection in both UD and DF colonoid monolayers transduced with MUC2 shRNA.
Importantly, monolayers are a suitable model to study host-microbial interactions at the apical surface of the epithelia. Colonoid monolayers form a thick attached MUC2-positive mucus layer upon differentiation which is not easily permeated by commensal E. coli HS bacteria (Figure 4), similar to what has been suggested in the normal human colon.19 However, enterohemorrhagic E. coli (EHEC), a human colonic pathogen, has been shown to have the ability to destroy the MUC2-enriched attached mucus layer to reach the apical surface of the epithelium (Figure 4).14,20 Remaining MUC2 is only present inside the goblet cells.
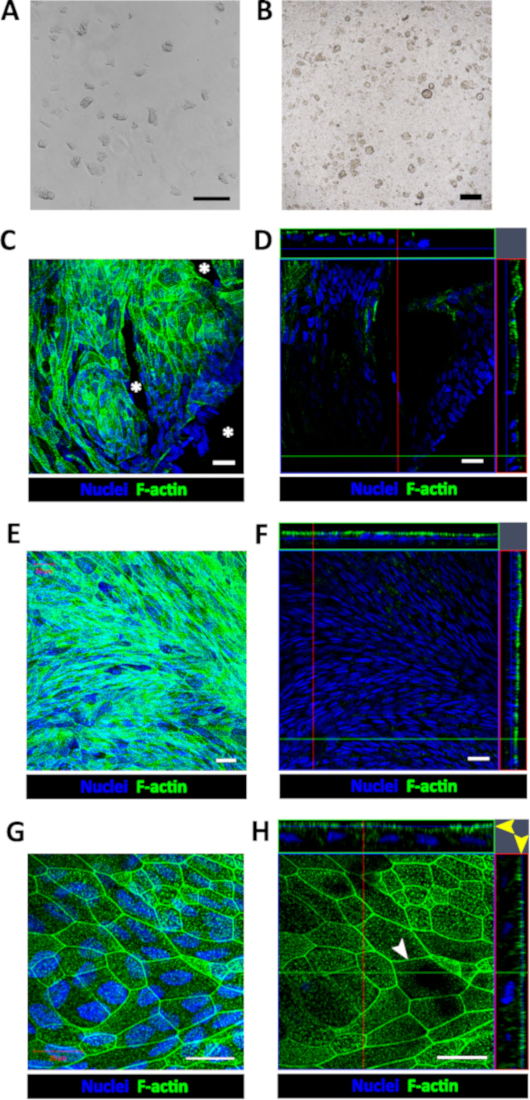
Figure 1: Establishment of human enteroid/colonoid monolayers. (A) Example of colonoid fragments after the dissolution of BMM and trituration. (B) Example of insert immediately after plating colonoid fragments. Scale bar (A-B) = 200 μm. Representative (C) maximum intensity projection and (D) confocal optical Z-section with the corresponding orthogonal projections show that colonoid fragments seeded onto human collagen IV-coated filters form multiple monolayer islands 2-4 days post-seeding. Cell-free areas (asterisk) are identifiable by the absence of both nuclear (Hoechst 33342, blue) and apical F-actin (phalloidin, green) staining in both C and D. Representative (E) maximum intensity projection and (F) confocal optical section with the corresponding orthogonal projections show a confluent colonoid monolayer with continuous apical surface detected by F-actin immunostaining approximately 1-week post-seeding. (G) High magnification of a representative maximum intensity projection and (H) confocal optical section with the corresponding orthogonal projections show that cells in confluent colonoid monolayers form the F-actin perijunctional rings (white arrowhead) and an immature apical brush border (yellow arrow heads). Scale bar (C-H) = 20 μm. Please click here to view a larger version of this figure.
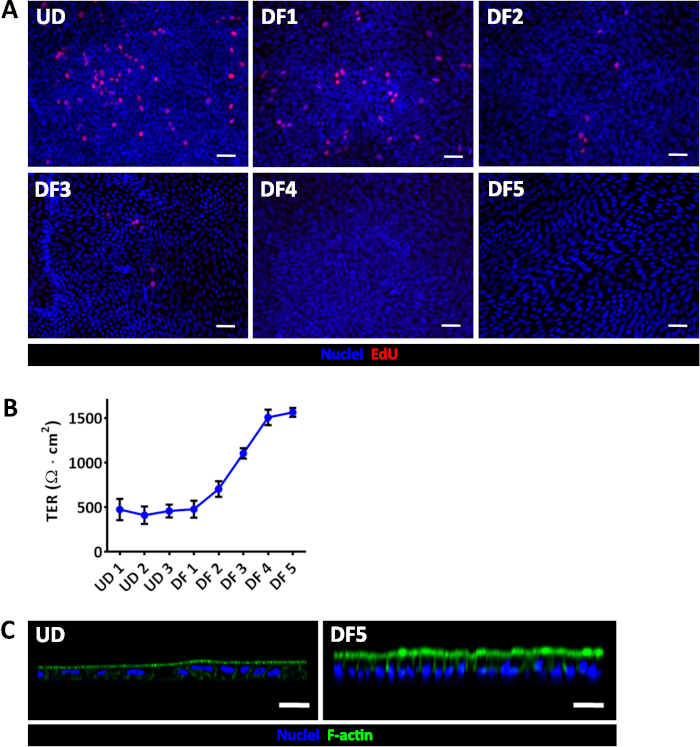
Figure 2: Evaluation of enteroid/colonoid monolayer differentiation. (A) EdU (red) incorporation demonstrates a progressive loss of proliferation during jejunal monolayer differentiation. (B) Average TER measurements in confluent jejunal monolayers in expansion or differentiation medium. Error bars represent SEM. UD, undifferentiated; DF, differentiated. Numbers correspond to days under the specified condition; UD1 was the first day of confluency, approximately 1 week after seeding. (C) UD jejunal monolayers have broad, shorter cells and a less-mature apical actin-based brush border than DF day 5 jejunal monolayers. Scale bar (A, C) = 50 μm. All monolayers were depicted at least 1-week post-seeding and were confluent prior to beginning differentiation. Please click here to view a larger version of this figure.
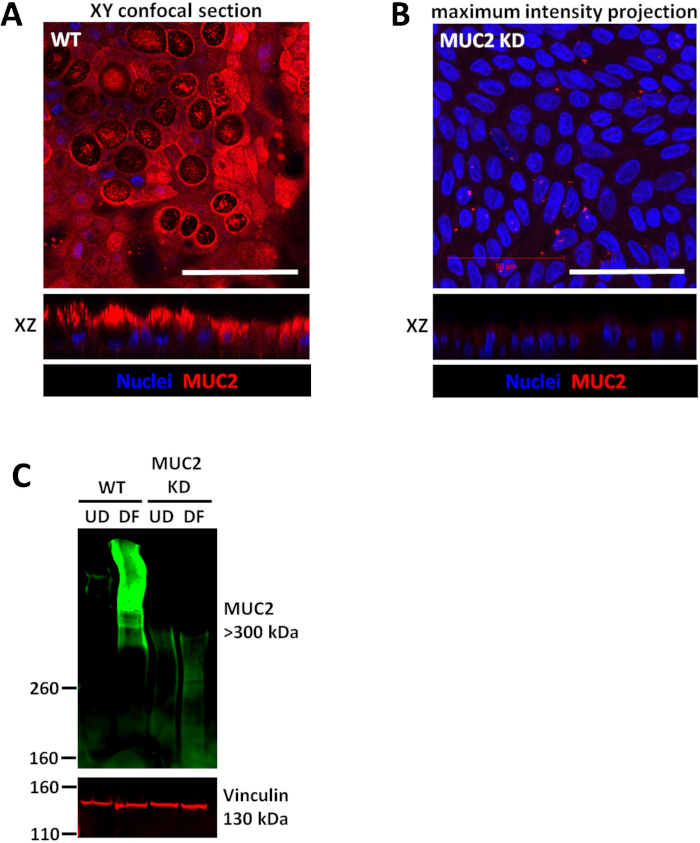
Figure 3: Wild type colonoids and shRNA transduced knockdown (KD) colonoids each form confluent monolayers. (A) Representative images of wild type (WT) human confluent colonoid monolayer differentiated for 5 days and (B) similarly grown monolayers derived from MUC2 KD colonoid cultures. Scale bar (A-B) = 50 μm (C) Representative immunoblot of colonoid cultures transduced with scrambled shRNA or MUC2 shRNA demonstrates that the changes in protein expression due to KD can be quantitated by immunoblot. Please click here to view a larger version of this figure.
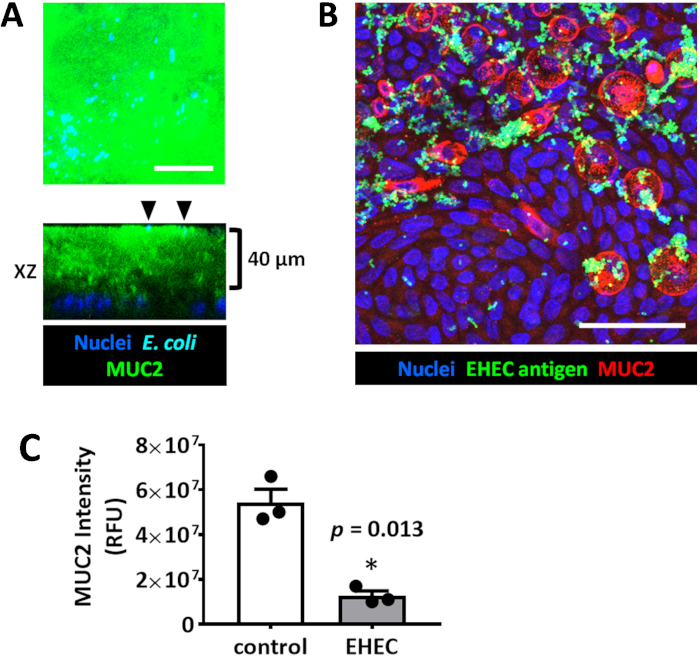
Figure 4: Enteroid/colonoid monolayers are suitable models to study luminal microbe-host interactions. (A) Representative maximum intensity projection and orthogonal optical section of a colonoid monolayer shows the thick apical MUC2-positive mucus layer which is not permeable to E. coli HS (black arrowheads) sitting at the apical mucus surface. The monolayer was infected for 6 hours with 106 cfu/mL HS. (B) Representative maximum intensity projection of human colonoid monolayer apically infected with EHEC (106 cfu/mL, 6 hours). Scale bar (A-B) = 50 μm. (C) MUC2 immunofluorescence intensity measurements show a significant decrease in MUC2 in EHEC-infected colonoid monolayers compared to uninfected controls. Error bars represent SEM. Please click here to view a larger version of this figure.
| Protein | Accession Number | Peptide Abundance |
| Basolateral Media | ||
| fatty acid-binding protein, liver [Homo sapiens] | NP_001434.1 | 1299 |
| profilin-1 [Homo sapiens] | NP_005013.1 | 797 |
| apolipoprotein A-IV precursor [Homo sapiens] | NP_000473.2 | 744 |
| ecto-ADP-ribosyltransferase 4 precursor [Homo sapiens] | NP_066549.2 | 633 |
| glyceraldehyde-3-phosphate dehydrogenase isoform 1 [Homo sapiens] | NP_002037.2 (+1) | 616 |
| serotransferrin precursor [Homo sapiens] | NP_001054.1 (+1) | 572 |
| vitamin D-binding protein isoform 3 precursor [Homo sapiens] | NP_001191236.1 (+2) | 567 |
| catalase [Homo sapiens] | NP_001743.1 | 427 |
| agrin isoform 1 precursor [Homo sapiens] | NP_940978.2 (+1) | 377 |
| lactotransferrin isoform 1 precursor [Homo sapiens] | NP_002334.2 (+1) | 262 |
| Apical Media | ||
| trefoil factor 3 precursor [Homo sapiens] | NP_003217.3 | 326 |
| trefoil factor 1 precursor [Homo sapiens] | NP_003216.1 | 304 |
| basement membrane-specific heparan sulfate proteoglycan core protein isoform a precursor [Homo sapiens] | NP_001278789.1 (+3) | 303 |
| filamin-B isoform 1 [Homo sapiens] | NP_001157789.1 (+2) | 240 |
| trefoil factor 2 precursor [Homo sapiens] | NP_005414.1 | 182 |
| deleted in malignant brain tumors 1 protein isoform b precursor [Homo sapiens] | NP_015568.2 (+3) | 169 |
| keratin, type II cytoskeletal 1 [Homo sapiens] | NP_006112.3 | 157 |
| myosin-9 [Homo sapiens] | NP_002464.1 | 150 |
| aminopeptidase N precursor [Homo sapiens] | NP_001141.2 (+1) | 145 |
| agrin precursor [Homo sapiens] | NP_940978.2 (+1) | 112 |
Table 1. A partial list of peptides identified by liquid chromatography/tandem mass spectrometry in apical and basolateral fluids sampled from differentiated jejunal monolayers.
