1. Detection of ROS burst in Arabidopsis leaf discs following immune elicitation
- Plant growth and maintenance.
- To synchronize germination, stratify Arabidopsis seeds by suspending approximately 50 seeds in 1 mL of sterile 0.1% agar [w/v] and store at 4 °C (no light) for 3-4 days.
NOTE: Stratify a wild type background control (for example, Col-0) and genotypes with high and low immune outputs (for example, cpk28-1 and bak1-5, respectively) to serve as internal controls. - Sow seeds on soil and germinate under standard short-day conditions (22 °C, 10 h light, 150 μE/m2/s light intensity, and 65-70% relative humidity).
- After approximately 7 days, use forceps to transplant individual seedlings to new pots, separated by at least 4 cm to permit full rosette development. Transplant 6-12 seedlings per genotype and return to standard short-day conditions. Regularly water and fertilize plants (1 g/L of 20-20-20 fertilizer every 2 weeks).
- To synchronize germination, stratify Arabidopsis seeds by suspending approximately 50 seeds in 1 mL of sterile 0.1% agar [w/v] and store at 4 °C (no light) for 3-4 days.
- Collect leaf discs in 96-well plates for overnight recovery.
- At 4-5 weeks post-germination (Figure 1A), use a sharp 4 mm biopsy punch to remove one leaf disc per plant, avoiding the mid-vein and being careful to limit wounding (Figure 1B). Place leaf discs in an unused 96-well luminometer plate containing 100 μL of ddH2O with the adaxial side facing upwards to prevent desiccation22 (Figure 1C). If assessing multiple elicitors, remove 1 leaf disc from the same leaf for each elicitor treatment.
NOTE: Sample leaves that are fully expanded, third to fifth from the top of the rosette, and approximately the same size and age to limit variability. If possible, cut leaf discs in half using a razor blade prior to overnight recovery, as this increases the surface area exposed to elicitor solution23. - Recover overnight at room temperature to prevent the interference of ROS produced by wounding during leaf disc collection. Cover plates with a lid to prevent evaporation.
- At 4-5 weeks post-germination (Figure 1A), use a sharp 4 mm biopsy punch to remove one leaf disc per plant, avoiding the mid-vein and being careful to limit wounding (Figure 1B). Place leaf discs in an unused 96-well luminometer plate containing 100 μL of ddH2O with the adaxial side facing upwards to prevent desiccation22 (Figure 1C). If assessing multiple elicitors, remove 1 leaf disc from the same leaf for each elicitor treatment.
- Perform the elicitor treatment and measure ROS production.
- Program the plate reader prior to adding the reaction solution, as this will reduce the time between ROS burst initiation and the first measurements recorded. Use a commercial software that is appropriate for the plate reader. In our case (see Indice), click Settings, and select the LUM96 Cartridge and the Kinetic Read Type. Click the Read Area category and drag to select a subset of wells or the entire plate to be read. Under the PMT and Optics tab, set the integration time to 1,000 ms. Under the Timing tab, set the Total Run Time to 40-60 min and the Interval to 2 min.
- Remove water from each well using a multi-channel pipette.
NOTE: Do not puncture leaf discs, as this may cause wounding stress and result in more variable ROS outputs. - Prepare a reaction solution containing 100 μM luminol, 10 μg/mL HRP, and the desired concentration of elicitor (for example: elf18 at 1 nM, 10 nM, 100 nM, or 1,000 nM) in sterile ddH2O using 10 mL of solution for one 96-well plate.
NOTE: Dissolve lyophilized peptides in sterile H2O in low-binding tubes to make a 10 mM stock, flash-freeze in liquid N2, and store at -80 °C. When ready to use, dilute stocks in sterile H2O to generate a 100 μM working stock and store at -20 °C. - Use a multi-channel pipette to dispense 100 μL of the reaction mixture to each well, adding the solution to all leaf discs of the same treatment at the same time22. Include a control reaction (no elicitor) for each genotype to assess basal ROS levels in the absence of elicitation. Immediately measure light emission for all wavelengths in the visible spectrum using a microplate reader.
NOTE: Prepare and apply the reaction solution under low light, as luminol and HRP are light-sensitive reagents. Keep reagents on ice or at -20°C unless in use. - Measure light emission with a 1,000 ms integration time in 2 min intervals over a 40-60 min period in a microplate reader in order to capture the dynamic oxidative burst (Figure 1D).
NOTE: Use a longer integration time to improve assay sensitivity11, while ensuring that all samples in a 96-well plate can be measured within one interval.
- Data interpretation.
- For each genotype, use a spreadsheet application to calculate the average photon count and standard error at each time point, and display ROS production as a function of time using a scatter plot.
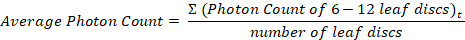
Where t = each time point - Alternatively, sum the photon counts for each leaf disc and present the average of these values for each genotype using a bar graph with standard error bars or a box and whisker plot.
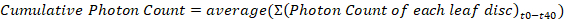
- For each genotype, use a spreadsheet application to calculate the average photon count and standard error at each time point, and display ROS production as a function of time using a scatter plot.
2. Seedling Growth Inhibition Assay
- Sterilize seeds and sow on Murashige and Skoog (MS) plates.
- Prepare sterile half-strength (0.5x) MS medium (2.16 g/L) containing 0.8% agar [w/v]. Pour media into plates (90 x 15 mm) in a laminar flow hood.
- Sterilize approximately 100 seeds in a microcentrifuge tube by washing with 1 mL of 70% ethanol for 2 min and remove by aspiration. Add 1 mL of 40% bleach and gently rock for 17 mins at room temperature. Remove bleach by aspiration and wash three times in 1 mL of sterile water for 5 min. Resuspend in 1 mL of sterile 0.1% agar.
NOTE: Alternatively, use a chlorine gas seed sterilization protocol24. - Sow approximately 100 seeds per genotype on MS agar plates using a pipette and seal with micropore tape in a laminar flow hood.
NOTE: Avoid placing seeds in close proximity as this will make transplanting seedlings difficult later. - Stratify seeds by placing plates at 4 °C (no light) for 3-4 days to synchronize germination (Figure 2A).
- Transplant seedlings into 48-well plates containing immune elicitor peptides.
- After stratification, move plates to light under short day conditions (22 °C, 10 h light, 150 μE/m2/s light intensity, and 65-70% relative humidity) for 3-4 days to allow germination.
- In a laminar flow hood, prepare elicitor peptide dilutions (0 nM, 1 nM, 10 nM, 100 nM, and 1,000 nM) in sterile 0.5x MS liquid media containing 1% sucrose [w/v], using 25 mL of MS for each 48-well plate. Prepare plates by pipetting 500 µL of MS liquid medium or MS medium containing peptides per well. If available, use a repeat pipettor for plate preparation to expedite the process.
NOTE: For each genotype, grow seedlings in MS without elicitor to account for any inherent differences in seedling growth. - Using sterile forceps carefully transplant one seedling to each well of the same size and age, ensuring that there is no damage to the seedling or breakage to the root and that the root is submerged in media. For each genotype, transplant a minimum of 6-8 seedlings into each peptide dilution (Figure 2B).
NOTE: Transplant seedlings at 4 days post-germination, as short roots are easier to manipulate, and older seedlings may yield less optimal results. - Seal plates with micropore tape and move back to light under standard short-day conditions (22 °C, 10 h light, 150 μE/m2/s light intensity, and 65-70% relative humidity). Allow seedlings to grow for 8-12 days.
- Determine percent growth inhibition.
- Carefully remove seedlings from 48-well plates and dry by dabbing on paper towel. Weigh seedlings on an analytical scale and record values. If available, use an analytical scale equipped with USB output to record fresh weight values on a spreadsheet. Before weighing seedlings, take a photo to visually display growth inhibition (Figure 2C).
- Determine percent growth inhibition of elicitor-treated seedlings compared to seedlings grown in MS only (Figure 2D) as follows:
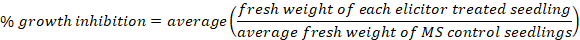
- Plot data using a bar graph with standard error bars or using a box and whisker plot to better display inter-experimental variance.
Mutant cpk28-119,25 and bak1-517,18 plants were used to demonstrate expected outcomes for genotypes with high and low immune responses, respectively, in oxidative burst and SGI assays relative to a wild-type background control (Col-0). To assess dose-dependent effects, a 10-fold peptide dilution series (1-1,000 nM) of elf18 was used. As expected, cpk28-1 loss-of-function lines had a higher cumulative (Figure 3A) and average (Figure 3B) ROS burst compared to Col-0, whereas bak1-5 displayed reduced ROS production at concentrations between 10 nM and 1,000 nM (Figure 3). Expected differences in SGI could be discerned between all genotypes grown in 100 nM and 1,000 nM elf18 (Figure 4A), which could also be visually observed in the 1,000 nM elf18 treatment (Figure 4B). Characteristic of high immune signaling, cpk28-1 mutants were markedly smaller than Col-0 when grown in 1,000 nM elf18, while bak1-5 mutants displayed weak growth inhibition relative to Col-0 due to disrupted MAMP detection.
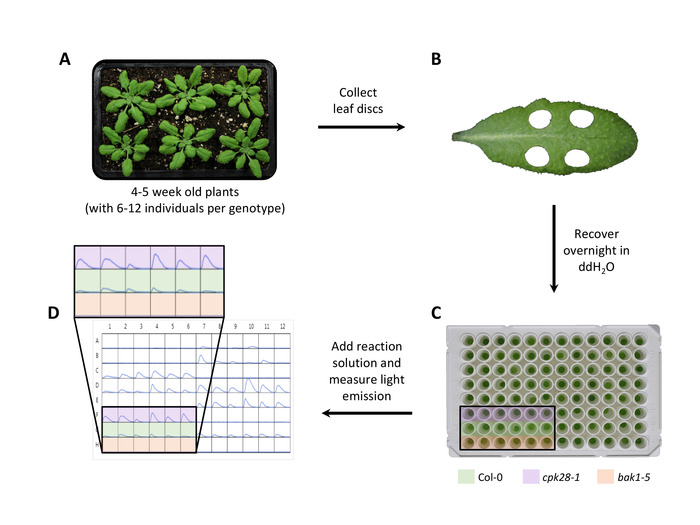
Figure 1. Luminol-based oxidative burst assay following immune induction in Arabidopsis. (A) Grow plants on soil in short day conditions for 4-5 weeks. (B) Use a 4 mm biopsy punch to collect leaf discs from each plant and recover overnight in ddH2O. (C) Add reaction solution (100 μM luminol, 10 μg/mL HRP, and the desired concentration of elicitor) and measure light emission over 40-60 min, in 2 min intervals with an integration time of 1,000 ms. (D) Determine average photon counts for each genotype relative to Col-0 (shown in green), a high ROS control such as cpk28-1 (shown in purple), and a low ROS control such as bak1-5 (shown in orange). Please click here to view a larger version of this figure.
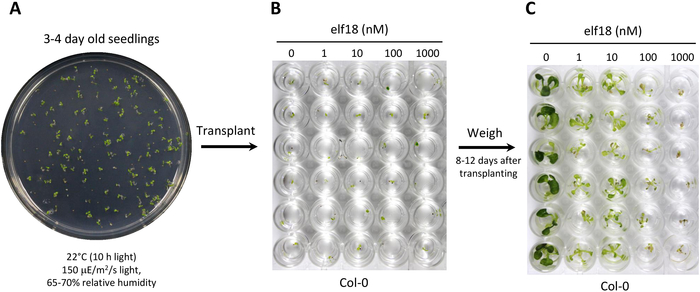
Figure 2. Elicitor-induced seedling growth inhibition assay in Arabidopsis. (A) Sow seedlings on MS agar and grow for 3-4 days under standard short-day conditions. (B) Transplant seedlings to 48-well plates containing MS medium or MS containing different concentrations of elf18. (C) After 8-12 days, visually assess the seedling size and then measure fresh weight using an analytical scale to determine percent growth inhibition. Please click here to view a larger version of this figure.
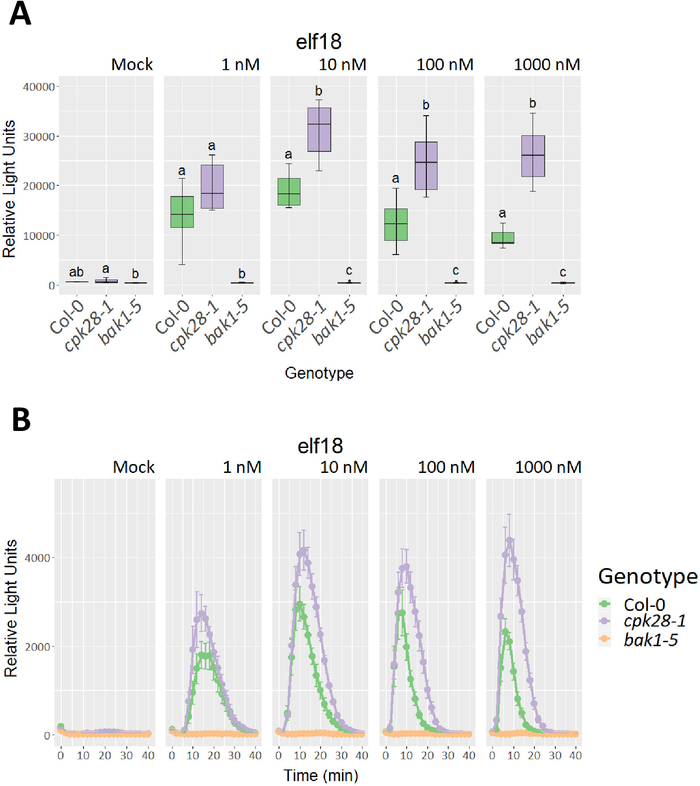
Figure 3. Representative elf18-induced oxidative burst in three Arabidopsis genotypes. Four-week-old plants were treated with a dilution series of elf18 (0 nM 'mock', 1 nM, 10 nM, 100 nM, and 1,000 nM). Col-0 represents the wild-type background control, with cpk28-1 and bak1-5 representing high and low ROS phenotypes, respectively. (A) Total photon count represented as relative light units following elf18 treatment (n=6 leaf discs from independent plants). Statistical differences are represented by lower-case letters and were calculated using a one-way ANOVA with a post-hoc Tukey's Honest Significant Difference test (p<0.05). (B) Average photon count, represented as relative light units, over 40 min following elf18 treatment (n=6 leaf discs from independent plants). Error bars represent standard error of the mean. Similar results were obtained in two of three experiments. Please click here to view a larger version of this figure.
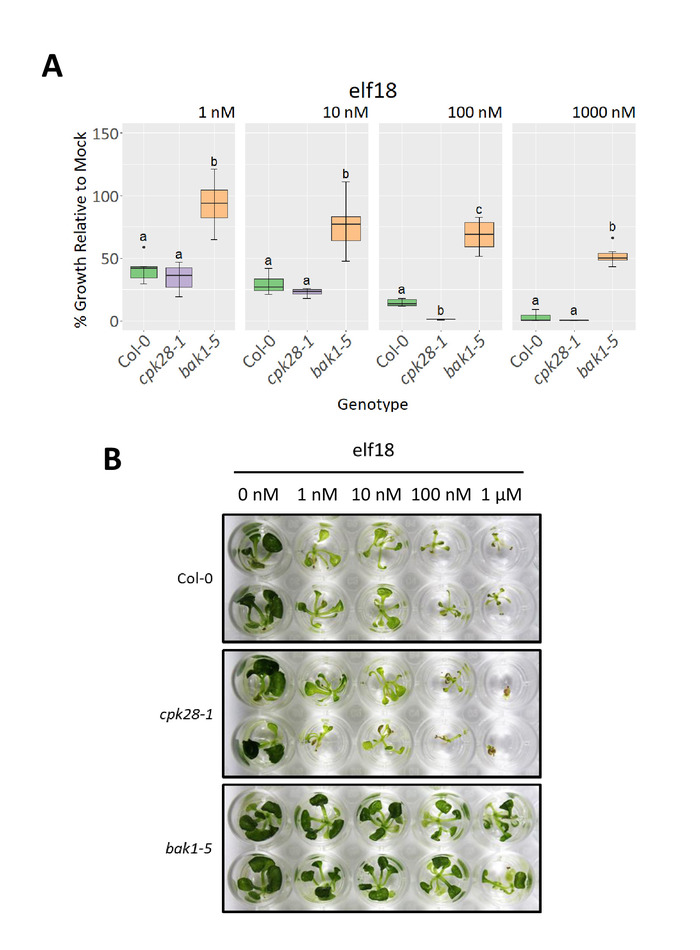
Figure 4. Representative elf18-induced seedling growth inhibition in three Arabidopsis genotypes. Wild-type (Col-0), cpk28-1, and bak1-5 seedlings were grown in a 10-fold dilution series (0-1,000 nM) of elf18. (A) Percent growth inhibition was calculated by comparing the weight of individual seedlings grown in elf18 (n=6 individual seedlings) to the average weight of seedlings of the same genotype grown in MS only ('mock') (n=6 individual seedlings) over a 10-day period. Statistical differences are represented by lower-case letters and were calculated using a one-way ANOVA with a post-hoc Tukey's Honest Significant Differences test (p<0.05). (B) Visual demonstration of SGI in two representative seedlings of Col-0, cpk28-1, and bak1-5 in response to increasing concentrations of elf18. Similar results were obtained in three of four experiments. Please click here to view a larger version of this figure.
