The main obstacle to obtaining adequate staining of Drosophila testes is the limited penetrability of antibodies, which has been partially resolved by using squashed preparations but at the cost of inducing a restriction of the 3D analyses (for example 3D representation or measure of colocalization). The procedure permits uniform labeling and preserves the volume of all the cells of the testis. Here we have focused the illustration of the method on the 3D representation of S4-S5 spermatocytes, as they are the largest cells and thus more sensitive to deformation from squashing. We used different nuclear markers, including nucleoporins and the labeling of the intranuclear MINT domain (Mad1-containing Intra Nuclear Territory) described by the mitotic protein Mad112. As MINTs could be revealed by several different antibodies, we used them to test the robustness of the protocol for quantitative 3D colocalization, a method that necessitates a correct spatial resolution.
The protocol described above permits the acquisition of confocal images of sufficient quality to then be treated for 3D reconstitution. Four adjacent serial sections of a stage S5 spermatocyte are depicted in Figure 1A, showing a representative immunostaining of MINT (Mad1 immunolabeling using GFP booster in red) together with DAPI (in cyan). The chromosomes are organized into the characteristic 3 major distinct clusters and the MINT structure is shown in fine detail around the DNA, intimately associated with–but distinct from–the autosomal chromatin masses. The resolution also permits detection of the presence of Mad1 at the nuclear periphery as well as between the autosomal masses of chromosomes 2 and 3. We used Imaris software for the 3D representation. The nuclear surface was defined by the low level presence of Mad1 at the nuclear envelope (in grey) and MINT was defined by high level accumulation of Mad1 inside the nucleus (in red) together with DAPI staining (in blue) (Figure 1A). The 3D reconstitution movie well illustrates the volume and the relationship of Mad1 (at the nuclear envelope and in the MINT) together with the chromosomes in stage S4–S5 spermatocytes.
The distribution of nucleoporins has been only poorly described in these cells, probably due to the difficulty detecting them in these large misshapen nuclei. The distribution of Nup154 has only been reported once previously16 and Nup153 and Nup98 were only studied in the spermatogonia and small, early spermatocytes17. The 3D reconstitutions of large S5 spermatocytes stained by the nucleoporin Nup62 and by importin β (product of the ketel gene) were feasible and are represented in Figure 1C and Figure 1D, respectively. These nuclear pore-associated components appear non-uniformly distributed over the nuclear rim as dots of irregular size. For example, the importin β gathers around the autosomal chromatin similarly to where a lamin network has been already described3. This kind of distribution with pore-rich areas and large pore-free areas has been described by electron microscopy in rodent spermatocytes18. Understanding the changes in lamin and nucleoporin distribution would help to understand the nuclear envelope structure in these G2 cells just prior to male meiosis.
While conserving the 3D volume of the cells this protocol also permits antibodies to penetrate uniformly into the depth of every cell, allowing quantitative analysis such as colocalization. The Pearson correlation coefficient (PCC) is a well-established metric. It can range from +1 (denoting perfect positive correlation) to −1 (perfect negative correlation)19. To evaluate the localization of MINT partner proteins relative to Mad1 in spermatocytes, we have previously estimated the PCC on a single section of 0.5 µm using ImageJ in S5 spermatocytes12 (Figure 2A,B). We tested the protocol more deeply by using 3D colocalization on the whole nuclei instead of just a surface. The volume of the nuclei was defined by manual segmentation and the PCC was measured using the Colocation tool. We analyzed eight S5 spermatocyte nuclei, from the same testis or different testes. Figure 2B shows a 3D representation of a nucleus that served to measure the PCC. Figure 2D shows the measurement of the PCC on eight different S5 spermatocyte nuclei from three different testes. PCC values are not different, varying from 0.6 and 0.75, indicating a high value for colocalization. These are indistinguishable from the PCC defined on a single section. Thus, the procedure permits a reproducible uniform labeling and preserves a good volume of all the cells of the testis.
Because the protocol allows reliable measurement of colocalization from independent testes, it permitted comparison of staining from different fly genotypes. This was illustrated here by comparison of WT flies and flies RNAi depleted for Mtor (generated by expression of dsRNA UAS lines and the Bam-Gal4 driver that achieve substantial depletion in late spermatogonia and early spermatocytes20). As described for the colocalization experiments, Mtor-RNAi-depleted testes and wild types testes were dissected and fixed together and the confocal images were acquired under constant settings. Respecting these conditions, the comparison of individual cells becomes possible. We observed that the MINT is more affected than the nuclear envelope by the loss of Mtor as shown by representative Mtor RNAi-depleted (upper) and wt (lower) cells (Figure 3)12.
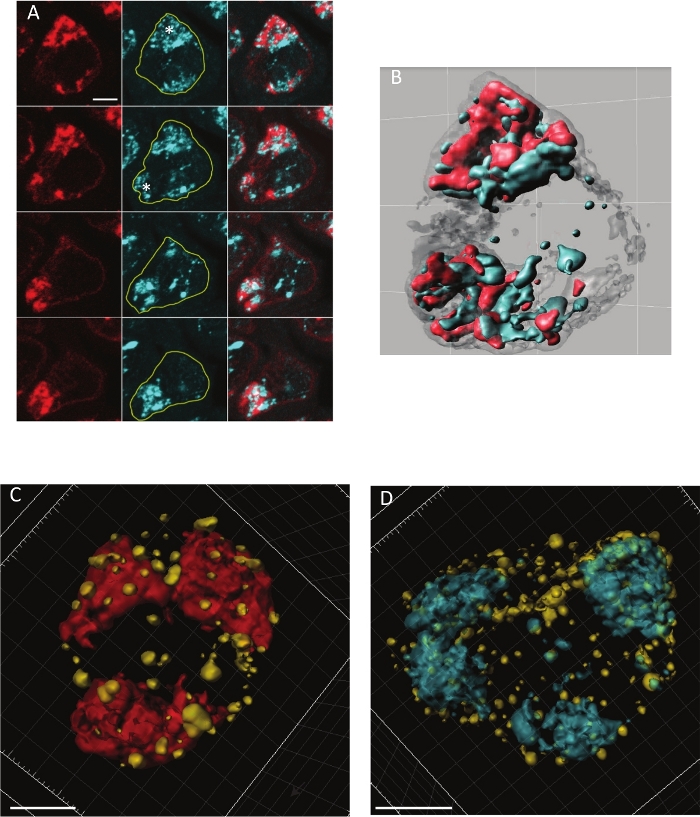
Figure 1: Illustration of immunostaining of Drosophila S4-S5 spermatocytes nuclei using different probes. (A) Images of a single stage 5 spermatocyte nucleus immunostained with α-GFP antibody (GFP booster) detecting the MINT structure by Mad1-GFP (red) together to DAPI staining (blue). It shows with precision the relationship between Mad1 and the two autosomal chromatin masses (represented as asterisks). Mad1 is also faintly detected at the periphery of the nucleus. All images are serial Z-projections of a stack of 3 (0.5 µm) successive confocal slices. Confocal images were acquired on a confocal microscope (63x, Plan Apochromatic oil DIC objective lens). Scale bars are 5 µm. (B) Frame from a movie rendering of the same spermatocyte nucleus. The nuclear surface was defined by low expression of Mad1 at the nuclear envelope (in greys) and MINT was defined by high expression of Mad1 inside the nucleus (in red). (A, B) Reprinted with permission from previously published figure (from12). (C) 3D image of a stage 5 spermatocyte nucleus with simultaneous immunostaining of Nup62 (yellow) and MINT (GFP booster in red). Scale bar is 4 µm. (D) 3D image of a stage 5 spermatocyte nucleus with simultaneous immunostaining of the importin-β Ketel (yellow) together with DNA (DAPI in blue). Scale bars are 5 µm. Please click here to view a larger version of this figure.
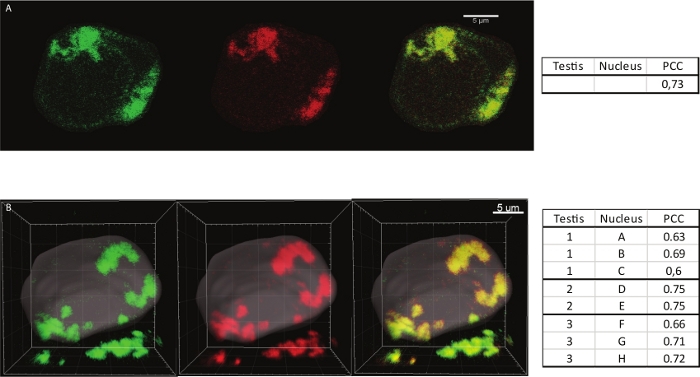
Figure 2: 2D and 3D Colocalization quantification of Mad1 together with Ulp1. (A) Merged images of a stage 5 spermatocyte nucleus immunostained with GFP booster antibody detecting Mad1 (green) and with anti-Ulp1 antibody (red) represented as a 1 µm thick stack of a spermatocyte nucleus. On the right is indicated the Pearson intensity correlation coefficient calculated using the Coloc-2 ImageJ plugin. The PCC was estimated to 0.73 (Reprinted with permission from Raich12). (B) Merged image of stage 5 spermatocyte nucleus (corresponding to nucleus G in the adjacent table) immunostained with GFP booster antibody detecting Mad1 (green) and with anti-Ulp1 antibody (red) represented in a 3D projection. The volume of the nucleus was defined by manual segmentation. On the right, the table lists the Pearson intensity correlation coefficients calculated on 8 different spermatocytes (A to H) from 3 different testes (1 to 3). Please click here to view a larger version of this figure.
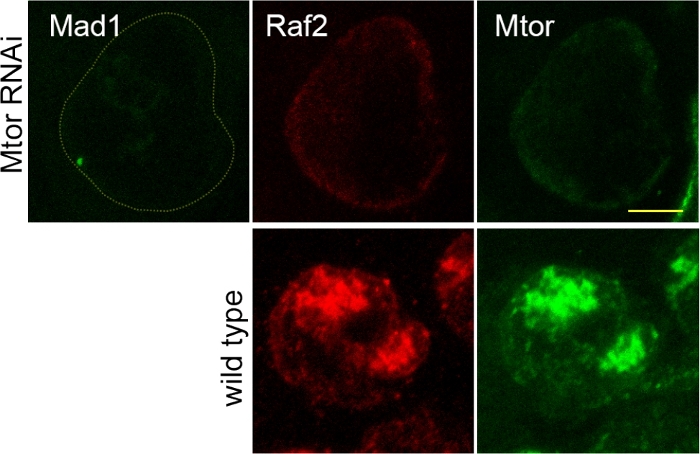
Figure 3: Immunofluorescence analysis of Mtor RNAi-treated spermatocytes compared to wild type. Cells depleted of Mtor using MtorRNAi (ID 110218) cells (upper panels) and wild type cells (lower panels) were stained for Mad1, Raf2 and Mtor proteins. The testes were fixed and stained together, mounted on the same slide and images acquired with the same parameters. The RNAi cells could be distinguished from WT by the absence or presence respectively of Mad1-GFP. The figure is representative of maximum projections of 4 x 0.5 µm stacks. Scale bar is 5 µm (Reprinted with permission from previously published figure in12). Please click here to view a larger version of this figure.
