The ability of this methodology to allow direct access to the intracellular cytosol depends on the spontaneous internalization of the SiNW into the cells. Although SiNWs will undergo spontaneous internalization into many cell types15, some cells, such as cardiomyocytes and neurons, will need the SiNWs to be treated to allow their internalization19. In this protocol we describe the internalization process of p-i-n SiNWs with 200-300 nm diameter and ~1-3 µm long into cardiac MFs. Figure 1A demonstrates how the SiNWs appeared under transmitted light microscopy. Using standard phase contrast optics, the confluent cells were easily apparent. However, the SiNWs were barely visible, making their locations impossible to define. Thus, we used dark field microscopy, where only reflecting objects can be seen and the background is dark. In this contrast, however, the cells cannot be seen, as they are not reflective. To show the location of the SiNW in the cells, we superimposed the images together, so that the perinuclear arrangement of the SiNWs within the cells is apparent. Although the colocalization of the SiNWs and cells is apparent, it still is important to verify that the SiNWs are indeed internalized within the cells, and not resting on the plasma membrane. To this end, we used confocal microscopy (Figure 1B), where the cytosol was stained with calcein-AM (green) and the plasma membrane with a membrane marker (red). The intracellular location of the SiNWs was then evident. Note that this step is extremely important in order to verify the intracellular location of the SiNWs, especially when a new cell type (or line) is used. However, after the internalization is established for the stained cells, other samples should be used for the stimulation procedure. Although it is not within the scope of this demonstration, it is also important to mention that in cases where SiNWs are not internalized, bioelectric processes may still be examined in a similar, extracellular way16. After the cells are hybridized with the SiNWs, they can be used to perform bioelectric studies.
Here we demonstrate the use of this methodology to compare the homo-cellular MF-MF electrical coupling to the hetero-cellular coupling of MFs to CMs. After the cells were hybridized with the SiNWs, the hybrids were seeded with CMs and cultured. It is important to limit the culture time so that the proliferating MFs do not overpopulate the culture. Figure 2 shows a representative example of such an experiment. Co-cultured cells were loaded with calcium sensitive dye (Fluo-4) and the cells were imaged under a spinning disc confocal microscope. Before applying any laser pulse, a brightfield image was obtained to identify the location of a SiNW that will be stimulated. Then, a short baseline video was recorded, so the spontaneously beating CMs and the resting MFs can be identified (Supplementary Video 1 and Supplementary Video 2). In this experiment, the identified SiNW was stimulated with a single point laser pulse (640 nm, 1 ms, 4 mW). The cells were constantly imaged before and after the stimulation to visualize the calcium dynamics upon optical stimulation. As cells in culture greatly differ in their brightness (Supplementary Video 1), the live videos were processed into dF/F videos with blue to red pseudocolor (Supplementary Video 2), so that the calcium signals will be clearer. In this method, each pixel is compared to its own baseline resting state, before the stimulation was applied. The representative images in Figure 2C demonstrate the calcium propagation within the MFs-CMs co-culture and can be further analyzed to study the inter-cellular electrical coupling between the different cells, as well as the intracellular calcium dynamics.
Figure 3A demonstrates a way to represent the calcium flux of the entire field of view in a single image. By analyzing the dF/F videos, we show the time at which the different cells were excited, which was determined by the point at which the change in average optical flow reaches its maximum. This way, one can see how the calcium flux propagated from cell to cell. A custom-built MATLAB script (Supplementary File 1) was written and used to calculate the optical flow and to aid in identifying the time of activation within each cell region (Figure 3B). The inter-cellular propagation speed can then be determined by measuring the distance between the centroids of each cell and dividing by the time difference in their activation. A similar analysis can be conducted that replaces distance with the number of junction crossings; for our purposes, the distance metric more faithfully presents the transmission speed as it also accounts for the time spent in intra-cellular propagation, which is non-negligible. Figure 3C demonstrates how the intra-cellular velocities were determined: for each cell, a line was drawn in the direction of calcium propagation and a kymograph was generated by plotting the intensity time profile along that line. We fit a line to the front of the activation wave in the kymograph, such that the slope of the line represented the inverse of the intra-cellular propagation speed. Figure 3D summarizes the different velocities (inter- and intra-cellular) for the different cells in the co-culture. All of the data represented in this graph were derived from the single video of a single representative field of view that was presented here.
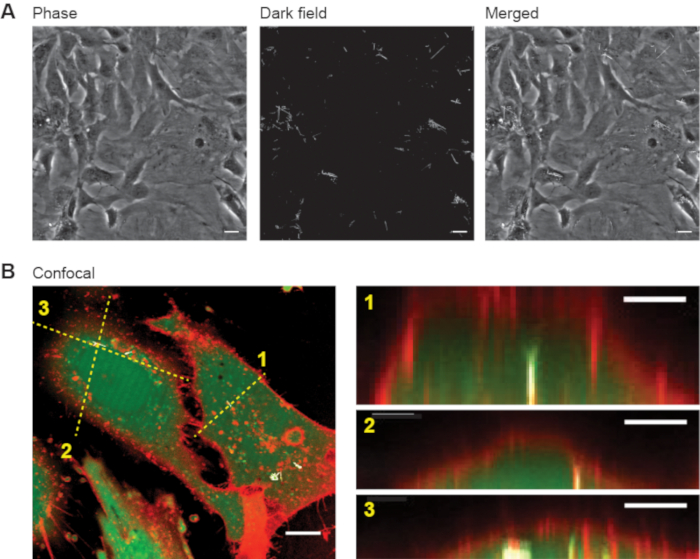
Figure 1: SiNWs internalized into MFs. (A) Phase contrast image of MFs treated with SiNWs show confluent cells (left). Darkfield image shows the SiNWs dispersed in the field of view (middle), while superimposing the two images (right) shows the perinuclear arrangement of the SiNWs within the MFs. Scale bars are 20 µm. (B) Representative confocal image (left, Scale bar is 10 µm) and 3 different n-z cross-sections that corresponds to the 3 dashed lines (right, Scale bars are 5 µm) show the inner part of a MF and the SiNW that are clearly inside the cytosol. The cytosol is green (Calcein-AM), the membrane red (cell mask), and the SiNWs white (reflection). Please click here to view a larger version of this figure.

Figure 2: Calcium propagation in MF-CM co-culture. (A) Illustration of a typical stimulation experiment. MFs with internalized SiNWs are stimulated with a laser pulse, while the calcium flux is monitored by a calcium sensitive dye. (B) A bright field image of the cells shows the internalized SiNW inside one of the MFs (left) and a representative fluorescence image of the calcium dye (right). The stimulated SiNW is highlighted by a pink arrowhead. (C) A co-culture of MFs and CMs was loaded with calcium sensitive dye and a SiNW was stimulated with a laser pulse (640 nm, 1 ms, 4 mW). The calcium flow was monitored by the Fluo-4 intensity (top images) and a dF/F video was derived from it (bottom images). The dF/F algorithm makes the visualization of the calcium flux clearer, as it compares the intensity of each pixel to its own baseline, thus showing the change, and not the actual intensity. Scale bars is 20 µm. Please click here to view a larger version of this figure.
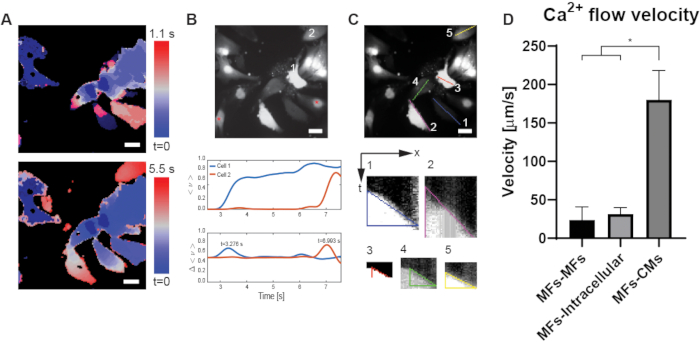
Figure 3: Analysis pf the dF/F videos from Figure 2. (A) the calcium propagation can be visualized via an optical mapping image, where the color codes for time the pixel was excited (meaning increase in calcium concentration and Fluo-4 relative intensity). The optical mapping can be done for different time frames, i.e. short (1.1 s, top image) or longer (5.5 s, bottom image) durations. Scale bars are 20 µm. (B) Representative examples for calculating cell-cell transmission speed. Both cells are MFs here, but the same computation can be performed with MF-CM transmission. The graphs show the traces for the average optical flow (middle) and the differential (bottom) for each cell. The cell-cell transmission can then be calculated using the Tmax for both cells as well as their centroids, as described in the text. Red asterisks denote two CMs with limited MF-CM coupling and hence lack of response to optical stimulation. Scale bars are 20 µm. (C) the MFs’ intracellular calcium flux velocity was derived from the slope of a kymograph generated by slicing the colored lines. The kymographs represent intensity along the line (x axis) and time (y axis), so that steeper slopes correspond to higher velocities. Scale bars are 20 µm. (D) The MFs-MFs and MFs-CMs inter-cellular velocities, as well as the MFs’ intra-cellular velocities can be derived from B and C respectively and plotted for comparison (p-value<0.0001). Please click here to view a larger version of this figure.
Supplementary Video 1: Effect of the MF-SiNW hybrid optical stimulation by Fluo-4. The CMs rate is illustrated before and after the stimulation for comparison. Please click here to view this video. (Right-click to download.)
Supplementary Video 2: Effect of the MF-SiNW hybrid optical stimulation by the dF/F video, derived from the Fluo-4 videos. The CMs rate is illustrated before and after the stimulation for comparison. Please click here to view this video. (Right-click to download.)
Supplementary Figure Please click here to download this figure.
