Preparation of Adipose Progenitor Cells from Mouse Epididymal Adipose Tissues
Summary
We present a simple method to isolate highly viable adipose progenitor cells from mouse epididymal fat pads using fluorescence activated cell sorting.
Abstract
Obesity and metabolic disorders such as diabetes, heart disease, and cancer, are all associated with dramatic adipose tissue remodeling. Tissue-resident adipose progenitor cells (APCs) play a key role in adipose tissue homeostasis and can contribute to the tissue pathology. The growing use of single cell analysis technologies – including single-cell RNA-sequencing and single-cell proteomics – is transforming the stem/progenitor cell field by permitting unprecedented resolution of individual cell expression changes within the context of population- or tissue-wide changes. In this article, we provide detailed protocols to dissect mouse epididymal adipose tissue, isolate single adipose tissue-derived cells, and perform fluorescence activated cell sorting (FACS) to enrich for viable Sca1+/CD31–/CD45–/Ter119– APCs. These protocols will allow investigators to prepare high quality APCs suitable for downstream analyses such as single cell RNA sequencing.
Introduction
Adipose tissue plays a key role in energy metabolism. Excess energy is stored in the form of lipids, and adipose tissue is capable of significant expansion or retraction depending on nutritional status and energetic demand. Expansion of adipose tissue can result from an increase in adipocyte size (hypertrophy) and/or from an increase in adipocyte number (hyperplasia); the latter process tightly regulated by proliferation and differentiation of adipose progenitor cells1,2. During obesity, adipose tissue excessively expands, and tissue dysfunction – including hypoxia, inflammation, and insulin resistance – often develops3,4. These complications are risk factors to many chronic diseases including hypertension, diabetes, cardiovascular diseases, stroke, and cancer5. Hence, limiting uncontrolled adipose tissue expansion and mitigating adipose tissue pathologies are top biomedical research priorities. During adipose tissue expansion, resident adipose tissue-derived stem cells (ASCs) proliferate and differentiate sequentially into preadipocytes (committed progenitor cells) and then into mature adipocytes6. Recent single-cell RNA-sequencing (scRNA-seq) studies show that these adipose progenitor cell (APC) populations (ASCs and preadipocytes) exhibit substantial molecular and functional heterogeneity7,8,9,10,11,12. For example, ASCs display a reduced adipogenic differentiation capacity, while also exhibiting higher proliferation and expansion capabilities, compared to preadipocytes7. Further molecular differences are reported within ASC and preadipocyte populations, although the functional relevance of these differences remains unclear7. Together, these data highlight the complexity of the adipose progenitor cell pool and underscore the need to develop and standardize tools to better understand and manipulate these critical cell populations.
This protocol details the isolation of high viability Sca1+ adipose progenitor cell populations from mouse epididymal fat pads that are suitable for sensitive downstream analyses, including single-cell studies (scRNA-sequencing) and cell culture. Isolation and dissociation of epididymal fat pads was performed as previously described7,13 with slight modifications that improve the viability of isolated APCs. In brief, dissociated cells from epididymal fat pads are stained with antibodies against Sca1, a marker for both ASCs and preadipocytes6,7, and other lineage (Lin) markers: Ter119 (erythroid cells), CD31 (endothelial cells), and CD45 (leukocytes). Viable Sca1+/Ter119–/CD31–/CD45–/DAPI– cells are then sorted by fluorescence activated cell sorting (FACS). Importantly, this protocol was validated by successful isolation and analysis of viable Sca1+/Lin– adipose progenitor cells reported in a recent single cell RNA sequencing study that identified functionally heterogeneous subpopulations within ASCs and preadipocytes7.
Protocol
All animal experimental procedures were performed under approval by the Mayo Clinic Institutional Animal Care and Use Committee.
1. Solution preparation
- Prepare collagenase 2% (w/v) solution by dissolving collagenase II 2 g in 100 mL Hanks' balanced salt solution (HBSS). Aliquot 200 µL each for each use.
- Prepare neutralization medium by mixing 84 mL F-12 medium, 15 mL horse serum, and 1 mL penicillin/streptomycin.
- Prepare flow cytometry buffer by dissolving 500 mg of bovine serum albumin (BSA) and 400 µL of 0.5 M EDTA in 100 mL of Dulbecco's phosphate-buffered saline (DPBS).
2. Dissection of epididymal fat pad and tissue dissociation
- Humanely euthanize (isoflurane/cervical dislocation) one male mouse. In this protocol, a four-month-old FVB mouse was used.
- Apply/spray 70% ethanol on the abdomen and expose the lower abdominal cavity using clean scissors and forceps.
- Locate epididymal fat pad (proximal/attached to testes). Hold the junction between testes and epididymal fat pad with blunt forceps and pull gently to liberate the epididymal fat pad.
- Remove testes by cutting using scissors and incubate epididymal fat pad in 5 mL of HBSS supplemented with 3% BSA in a 50 mL conical tube at room temperature for 15 min.
- Centrifuge at 150 x g for 7 min at 4 °C.
- Take floating epididymal fat pad from the 50 mL conical tube and finely mince the tissue using clean scissors.
- Add 200 µL collagenase 2% solution into 3.8 mL of HBSS in a 13 mL culture tube. Add the minced tissue and incubate in a rotating incubator at 5 rpm for 1 h at 37 °C. Instead of a rotating incubator, cell culture incubator at 37 °C can be alternatively used with occasional agitations.
- Transfer entire contents into a 50 mL conical tube and add 10 mL of neutralization medium. Mix gently by inverting the tube 2-3 times.
- Prepare another 50 mL conical tube fitted with a 70 µm cell strainer. Filter the digested tissue into the new tube using the cell strainer.
- Transfer the flow-through to a 15 mL conical tube and centrifuge at 350 x g for 10 min at 4 °C.
- Carefully remove the supernatant and resuspend cell pellets with 5 mL of DPBS. Centrifuge at 350 x g for 10 min at 4 °C.
- Carefully remove the supernatant and add 50 µL of flow cytometry buffer. Mix well by pipetting gently and keep cells on ice. Due to the volume of the cell pellets and small amount of residual supernatant, total volume of cells in the flow cytometry buffer will be greater than 50 µL (approximately 100 µL).
3. Antibody labeling and fluorescence activated cell sorting (FACS)
- After mixing the cells well with gentle pipetting, transfer 54 µL of the cell suspension into a 1.7 mL microcentrifuge tube. Add 6 µL of FcR blocking reagent and mix well by pipetting gently. Incubate at 4 °C for 10 min. Save any leftover cells after taking 54 µL and keep them in the ice to use as an unstained control in step 3.5 and step 3.6.
- During the incubation in step 3.1, prepare an antibody cocktail by combining 11 µL each of anti-Sca1-APC, anti-Ter119-FITC, anti-CD31-FITC, and anti-CD45-FITC antibodies in a microcentrifuge tube. Mix well by gentle pipetting and keep on ice protected from light.
- After incubation with FcR blocking reagent for 10 min, add 40 µL of the antibody cocktail and mix well by gentle pipetting. Incubate at 4 °C for 10 min.
- Add 500 µL of flow cytometry buffer supplemented with 1 µg/mL of DAPI into the stained cells. Mix well by gentle pipetting and filter cells using a 5 mL test tube with cell strainer snap cap. Keep cells on ice.
- Add 500 µL of flow cytometry buffer to remaining cells in step 3.1 and mix well by gentle pipetting. Filter cells using a 5 mL test tube with cell strainer snap cap. Keep these cells on ice and use as an unstained control in the following step.
- Take the stained cells and unstained control to FACS analysis or sorting instrument.
- Identify and isolate the APC+/FITC–/DAPI– population with gating strategies shown in Figure 1. These cells represent viable Sca1+/Ter119–/CD31–/CD45– cells. Debris and cell aggregates are depleted using forward (FSC) and side scatter (SSC) plots. Then, DAPI– population is gated, followed by gating of APC+/FITC– population. Unstained controls should be used to aid in setting gating parameters.
- Collect the cells into 500 µL of flow cytometry buffer in a 1.5 mL microcentrifuge tube. Moving forward, all procedures should be performed under sterile conditions to minimize a contamination. Approximately 50,000 – 100,000 cells are collected from one mouse.
- Count cells using a hemocytometer to evaluate cell viability. Since cell aggregates can interfere with further downstream analyses, the presence of cell aggregates is also evaluated to ensure a minimal presence (<5% of viable cell number) of these aggregates.
Representative Results
Four-month-old male FVB mice were used in this experiment. After exclusion of debris and doublets using FSC/SSC plots, viable cells (DAPI– population) were gated, followed by the selection of APC+/FITC– population (Figure 1). DAPI, APC, and FITC gates were drawn based on the unstained control. Gating strategies are shown in Figure 1.
After 1 h of sorting, the quality of isolation was quantitatively evaluated by flow cytometry analysis (Figures 2,3). The cells were stained with propidium iodide solution (1:100) for viability staining. Using the same gating for sorting, the cells maintained high viability and purity: 92.6 ± 2.2% single cells (third panel in Figure 2), 86.4 ± 3.0% viability (PI– cells), and 86.0 ± 2.8% APC+/FITC– cells (n= 4, average ± standard deviation). These percentages were defined as percentages of each gated population (single cells, PI– cells, and PI–/APC+/FITC– population, respectively, in Figure 2) relative to total cell number.
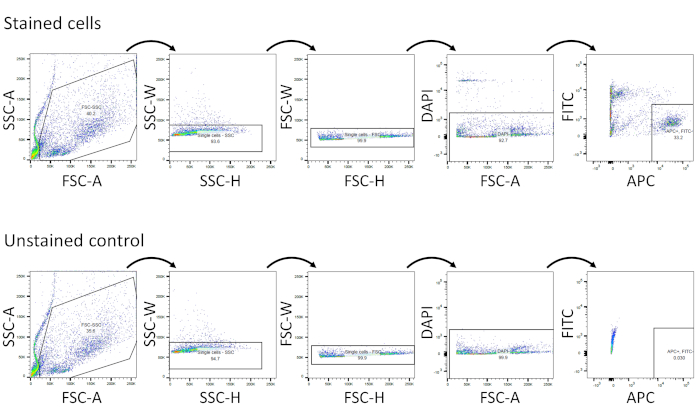
Figure 1: Isolation of viable Sca1+ adipose progenitor single cells by FACS. Gating strategies are shown in both stained cells and in an unstained control. Please click here to view a larger version of this figure.
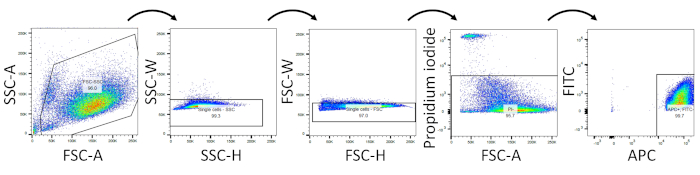
Figure 2: Representative flow cytometry analysis to assess post-isolation cell viability. Viability staining was performed using propidium iodide. Please click here to view a larger version of this figure.
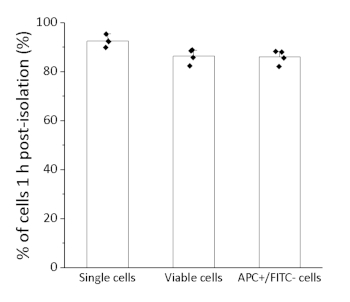
Figure 3: Quantification of percentage of cells 1 h post-isolation in flow cytometry analysis. Percentages of single cells, viable cells, and APC+/FITC– cells were quantified to evaluate purity and viability after isolation of cells. Data shown represents the average ± standard deviation. n=4. Please click here to view a larger version of this figure.
Discussion
Single cell RNA sequencing (scRNA-seq) is rapidly gaining traction as a powerful tool to simultaneously study diverse cell populations at the single cell level. Due to high costs associated with sample preparation and high throughput sequencing, it is imperative to optimize cellular inputs (high viability and purity) to increase the likelihood of experimental success. Some cell preparation protocols rely on removal of dead cells and debris using low-spin washes and column-based separation without FACS sorting14. Many of these methods, however, significantly reduce the number of viable recovered cells, and in many cases, dead cells are not completely removed14. This protocol outlines a simple method to isolate highly viable Sca1+ adipose progenitor cells containing minimal cellular debris, cell doublets, and dead cells. Although cells isolated using this protocol exhibited high viability, we still recommend minimizing time between cell isolation and further downstream analysis to maintain high viability. In addition, successful antibody labeling is critical to isolate Sca1+ adipose progenitor cells with high purity. Hence, determination of optimal concentration of antibodies is strongly recommended for each antibody, and dilution of the antibodies in step 3.2 can be adjusted accordingly.
This flow cytometry-based protocol is amenable to customization depending on individual interest in specific adipose progenitor cell sub-populations. Here, alternative antibody combinations can be used to identify and isolate the population(s) of interest. Indeed, due to extensive APC marker diversity, multiple labs studying APCs using scRNA-seq report isolating cells using other surface markers. For example, PDGFRA+/CD44+ cells, CD31–/CD45–/Ter119–/CD29+/CD34+/Sca1+ cells, and Pdgfrb+ cells have been isolated by FACS for downstream APC scRNA-seq analyses8,9,11. CD142+ subpopulations have also been characterized and are shown to exhibit limited adipogenic differentiation capacity and an ability to suppress adipogenesis10,11. In addition to Sca1, combinations of antibodies against CD55, CD81, and CD9 were recently used to prospectively isolate ASC and preadipocyte subpopulations (using this protocol) and study subpopulation molecular and functional heterogeneity7. Moreover, since adipose progenitor cells from other fat pads including inguinal fat pads and brown adipose tissues have been isolated using collagenase dissociation like the present protocol15,16,17, the present protocol may be applicable to isolation of adipose progenitor cells from other adipose tissues.
One caveat of the current approach is that it is tailored towards the isolation of immature APCs. Although the present protocol yields high viability APCs, the efficacy and efficiency of isolation of other adipose tissue resident cell types (i.e., immune cells, fibroblasts, endothelial cells, etc.) is unclear. Importantly, this protocol is not suitable for the isolation of mature adipocytes. Adipocytes have been extremely difficult to isolate using FACS due to their large cell size and fragility. Recently, isolation of mature adipocytes by FACS was reported using a large nozzle size (150 µm) and low sheath pressure (6 psi) with a specific FSC/SSC gating strategy18. Future studies could perhaps merge this adipocyte isolation protocol with the protocol described here to establish a more comprehensive pipeline to isolate and study diverse cell types within adipose tissue.
Divulgazioni
The authors have nothing to disclose.
Acknowledgements
We acknowledge the Mayo Clinic Microscopy Cell Analysis Core Flow Cytometry Facility for assistance with FACS sorting.
Materials
| 1.7 mL microcentrifuge tube | VWR | 87003-294 | |
| 13 mL culture tube | Thermo Fisher Scientific | 50-809-216 | |
| 15 mL conical tube | Greiner Bio-one | 188 271 | |
| 5 mL test tube with cell strainer snap cap | Thermo Fisher Scientific | 08-771-23 | |
| 50 mL conical tube | Greiner Bio-one | 227 261 | |
| 70 µm cell strainer | Thermo Fisher Scientific | 22-363-548 | |
| Anti-CD31-FITC antibody | Miltenyi Biotec | 130-102-519 | |
| Anti-CD45-FITC antibody | Miltenyi Biotec | 130-102-491 | |
| Anti-Sca1-APC antibody | Miltenyi Biotec | 130-102-833 | |
| Anti-Ter119-FITC antibody | Miltenyi Biotec | 130-112-908 | |
| BSA | Gold Biotechnology | A-420-500 | |
| Collagenase type II | Thermo Fisher Scientific | 17101-015 | |
| DAPI | Thermo Fisher | D1306 | |
| Dulbecco's phosphate-buffered saline (DPBS) | Thermo Fisher Scientific | 14190-144 | |
| F-12 medium | Thermo Fisher Scientific | 11765-054 | |
| FcR blocking reagent | Miltenyi Biotec | 130-092-575 | |
| Hanks' balanced salt solution (HBSS) | Thermo Fisher Scientific | 14025-092 | |
| Horse serum | Thermo Fisher Scientific | 16050-122 | |
| Penicillin-streptomycin | Thermo Fisher Scientific | 15140-122 | |
| Propidium iodide solution | Miltenyi Biotec | 130-093-233 |
Riferimenti
- Krotkiewski, M., Bjorntorp, P., Sjostrom, L., Smith, U. Impact of obesity on metabolism in men and women. Importance of regional adipose tissue distribution. The Journal of Clinical Investigation. 72, 1150-1162 (1983).
- Salans, L. B., Horton, E. S., Sims, E. A. Experimental obesity in man: cellular character of the adipose tissue. The Journal of Clinical Investigation. 50, 1005-1011 (1971).
- Trayhurn, P. Hypoxia and adipose tissue function and dysfunction in obesity. Physiological Reviews. 93, 1-21 (2013).
- Halberg, N., et al. Hypoxia-inducible factor 1alpha induces fibrosis and insulin resistance in white adipose tissue. Molecular and Cellular Biology. 29, 4467-4483 (2009).
- Upadhyay, J., Farr, O., Perakakis, N., Ghaly, W., Mantzoros, C. Obesity as a Disease. Medical Clinics of North America. 102, 13-33 (2018).
- Cawthorn, W. P., Scheller, E. L., MacDougald, O. A. Adipose tissue stem cells meet preadipocyte commitment: going back to the future. Journal of Lipid Research. 53, 227-246 (2012).
- Cho, D. S., Lee, B., Doles, J. D. Refining the adipose progenitor cell landscape in healthy and obese visceral adipose tissue using single-cell gene expression profiling. Life Science Alliance. 2, 201900561 (2019).
- Burl, R. B., et al. Deconstructing Adipogenesis Induced by beta3-Adrenergic Receptor Activation with Single-Cell Expression Profiling. Cell Metabolism. 28, 300-309 (2018).
- Hepler, C., et al. Identification of functionally distinct fibro-inflammatory and adipogenic stromal subpopulations in visceral adipose tissue of adult mice. eLife. 7, 39636 (2018).
- Merrick, D., et al. Identification of a mesenchymal progenitor cell hierarchy in adipose tissue. Science. 364, 2501 (2019).
- Schwalie, P. C., et al. A stromal cell population that inhibits adipogenesis in mammalian fat depots. Nature. 559, 103-108 (2018).
- Raajendiran, A., et al. Identification of Metabolically Distinct Adipocyte Progenitor Cells in Human Adipose Tissues. Cell Reports. 27, 1528-1540 (2019).
- De Matteis, R., et al. In vivo physiological transdifferentiation of adult adipose cells. Stem Cells. 27, 2761-2768 (2009).
- Hanamsagar, R., et al. An optimized workflow for single-cell transcriptomics and repertoire profiling of purified lymphocytes from clinical samples. Scientific Reports. 10, 2219 (2020).
- Marinovic, M. P., et al. Crotamine induces browning of adipose tissue and increases energy expenditure in mice. Scientific Reports. 8, 5057 (2018).
- Takahashi, H., et al. Biological and clinical availability of adipose-derived stem cells for pelvic dead space repair. Stem Cells Translational Medicine. 1, 803-810 (2012).
- Cowan, C. M., et al. Adipose-derived adult stromal cells heal critical-size mouse calvarial defects. Nature Biotechnology. 22 (5), 560-567 (2004).
- Hagberg, C. E., et al. Flow Cytometry of Mouse and Human Adipocytes for the Analysis of Browning and Cellular Heterogeneity. Cell Reports. 24, 2746-2756 (2018).
