Elevated resistance to FSS-induced mechanical destruction has been previously shown to be a conserved phenotype across multiple cancer cell lines and cancer cells freshly isolated from tumors relative to non-transformed epithelial cell comparators15,24. Here, additional cancer cell lines from a variety of tissue origins (Table 2) were tested to demonstrate that the majority of these cells display viability ≥ 20% after 10 pulses of FSS at 250 µL/s. The one exception is MiaPaCa2 cells, which were relatively sensitive to mechanical destruction from FSS (viability ≤ 10%). To adequately describe the FSS resistance profile of a cell line, n ≥ 3 biological replicates are recommended.
By way of comparison, all of the non-transformed epithelial cells examined have viability < 10% under these conditions15,24. Thus, while there is a range in FSS resistance observed, the majority of the cancer cell lines tested exhibit greater FSS resistance than non-transformed cells. Cancer cell lines can be derived from both primary tumor tissues and metastases. One could postulate that cells derived from metastases may exhibit greater FSS resistance as this phenotype may have been selected during metastatic dissemination. However, the FSS resistance level was shown to not depend on whether cells were derived from primary tumors or metastases15,24. Moreover, the levels of FSS resistance did not correlate with metastatic potential in a series of human prostate cancer cell lines15.
To test this further, BALB/c mammary epithelial cells with varying metastatic potential (4T1 = highly metastatic, 4T07 = weak to moderate metastatic potential, 67NR = no to low metastatic potential25,26) were used. This experiment revealed that FSS resistance is not correlated with metastatic potential (Figure 1). Moreover, both 4T1 and 4T07 cells exhibit a biphasic loss of cell viability-a greater loss of viability in pulses 1-2 than observed in subsequent pulses. This is typical of most cancer cell lines investigated by this group. In contrast, 67NR exhibits a more linear loss of cell viability as a function of FSS. Collectively, the data from Table 2 and Figure 1 demonstrate that FSS resistance is a property of transformed cells.
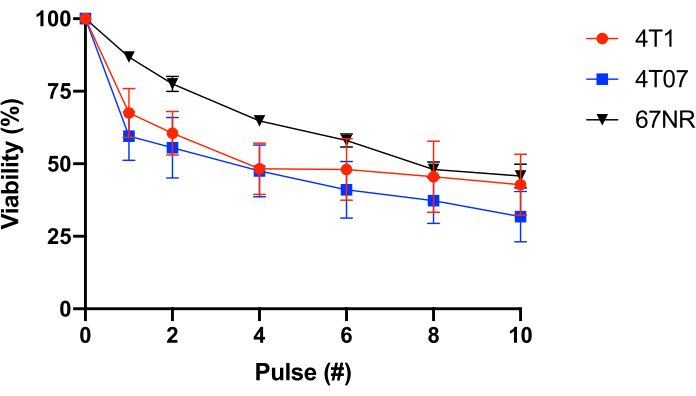
Figure 1: Fluid shear stress resistance of syngeneic BALB/c mammary epithelial cancer cells. Cells were exposed to FSS (30 G needle, 10 pulses@250 mL/s), and viability was measured using resazurin conversion (n = 4/cell line). While FSS exposure reduced the number of viable cells (p < 0.0001, 2-way ANOVA), and each cell line displayed different resistance profiles (p = 0.0446, 2-way ANOVA), there was no significant difference among cell lines after 10 pulses of FSS exposure (p = 0.2833, 2-way ANOVA). Abbreviations: FSS = fluid shear stress; ANOVA = analysis of variance. Please click here to view a larger version of this figure.
| Shear (τ): | wall (maximum) | minimum | |||||||||||
| Cell Diameter: | N/A | 10 µm | 15 µm | 20 µm | |||||||||
| Needle Gauge: | 30 | 27 | 25 | 30 | 27 | 25 | 30 | 27 | 25 | 30 | 27 | 25 | |
| Flow Rate (µL/s) | 20 | 507 | 220 | 116 | 32 | 10 | 4 | 48 | 16 | 7 | 64 | 21 | 9 |
| 50 | 1267 | 550 | 290 | 80 | 26 | 11 | 120 | 39 | 17 | 159 | 52 | 22 | |
| 100 | 2534 | 1100 | 580 | 159 | 52 | 22 | 239 | 79 | 33 | 319 | 105 | 45 | |
| 150 | 3801 | 1650 | 869 | 239 | 79 | 33 | 359 | 118 | 50 | 478 | 157 | 67 | |
| 200 | 5068 | 2200 | 1159 | 319 | 105 | 45 | 478 | 157 | 67 | 637 | 210 | 89 | |
| 250 | 6335 | 2750 | 1449 | 398 | 131 | 56 | 598 | 196 | 84 | 797 | 262 | 111 | |
Table 1: Maximum shear stress (τwall) levels. The table lists the maximum wall FSS levels in dyn/cm2 for 30 G, 27 G, and 25 G needles at the flow rates of 20, 50, 100, 150, 200, and 250 µL/s. Shear stress levels were calculated using the Poiseuille equation (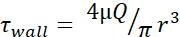 ), available information for the inner diameter of each needle gauge, as well as the assumption that µ = 0.01 dyn·s/cm2. Minimum FSS levels for each size were calculated using
), available information for the inner diameter of each needle gauge, as well as the assumption that µ = 0.01 dyn·s/cm2. Minimum FSS levels for each size were calculated using 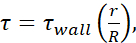 wherein r is the radius of cell, and R is the radius of the needle. Abbreviation: FSS = fluid shear stress; τ = shear; τwall = maximum shear; µ = viscosity; Q = volumetric flow rate.
wherein r is the radius of cell, and R is the radius of the needle. Abbreviation: FSS = fluid shear stress; τ = shear; τwall = maximum shear; µ = viscosity; Q = volumetric flow rate.
| Cell Line | Tissue Source | Species | Mean Viability (%) after 10 pulses |
| TRAMPC1 | Prostate | Mouse | 40 |
| 4T01 | Breast | Mouse | 32 |
| 4T7 | Breast | Mouse | 43 |
| 67NR | Breast | Mouse | 46 |
| 66CL4 | Breast | Mouse | 28 |
| RT4 | Bladder | Human | 62 |
| W17-266-4 | Melanoma | Human | 46 |
| HS852 | Melanoma | Human | 41 |
| HS695 | Melanoma | Human | 41 |
| A2058 | Melanoma | Human | 37 |
| A375 | Melanoma | Human | 37 |
| RPMI-7951 | Melanoma | Human | 35 |
| SKMEL2 | Melanoma | Human | 29 |
| A101D | Melanoma | Human | 28 |
| MiaPaCa | Pancreatic | Human | 7 |
Table 2: Fluid shear stress resistance of various cancer cell lines. Each cancer cell line was exposed to fluid shear stress from the syringe and needle model (30 G needle, 10 pulses@250 mL/s) (n ≥ 3/cell line), and viability was measured either by luciferase activity or resazurin conversion.
