Frozen offspring heart tissues were received from Dr. Sean Newcomer along with the institutional IACUC approval letter. The heart tissues were obtained from a long-term longitudinal study, flash-frozen in liquid nitrogen, and stored at -80 °C for future use. All protocols concerning the processing of offspring heart tissue followed the guidelines of Kansas City University IBC and IACUC committees.
1. Preparation of buffers and reagents
NOTE: Prepare all samples as per the manufacturer's guidelines. Use ultra-purified water or equivalent in all recipes and protocol steps. Wear personal protection equipment (lab gloats, facemask, gloves, and goggles/face shield) during this procedure. Buffer volumes are suitable for processing six tissue samples.
- Prepare 1.5 L tissue washing buffer by combining 154.035 g of sucrose (0.3 M final), 3.904 g of HEPES (10 mM final), 0.111 g of EDTA (0.2 mM final). Add ingredients to 1.0 L of water while on the stir plate, adjust the pH to 7.2 with dilute HCl, and makeup to final volume to 1.5 L.
NOTE: Approximately 200 mL of washing buffer is needed per sample (50-300 mg). Six tissue samples can be processed in a day. Use the remaining washing buffer to make an isolation buffer. - Prepare the isolation buffer from the wash buffer previously described in step 1.1 by adding 210 mg of BSA in 210 mL of wash buffer. Adjust the pH to 7.4 with dilute NaOH solution.
NOTE: Approximately 35 mL of isolation buffer is needed per tissue sample; therefore, 210 mL of isolation buffer is required for six samples. Prepare this buffer by adding fresh BSA solution (1mg/mL final) on the day of preparation. BSA is used to isolate mitochondria as it improves the mitochondrial properties by removing natural uncouplers such as free fatty acids. BSA has been found to have antioxidant activity by increasing oxidation rate of substrates, especially succinate, in brain mitochondria7. Alternatively, bovine serum albumin can be substituted by less expensive acid-free analogs such as ovalbumin. - Prepare 150 mL of resuspension buffer by combining 0.011 g of EDTA (final concentration of 0.2 mM EDTA or use 150 µL of 200 mM stock EDTA solution for 150 mL of resuspension buffer), 12.836 g of sucrose (0.25 M sucrose final concentration), and 0.236 g of Tris (10 mM Tris-HCl final). Adjust the pH to 7.8 with 0.1 N NaOH. Add 60 µL of the protease inhibitor cocktail just before use.
- Prepare the trypsin solution by dissolving 7.5 mg of trypsin in 3.0 mL of 1 mM HCl (2.5 mg of trypsin/mL final). This amount is sufficient for one sample.
NOTE: Prepare the trypsin solution based on the number of tissue samples planned to be processed. - Prepare the trypsin inhibitor solution from soybean by combining 13.0 mg of trypsin inhibitor in 20 mL of isolation buffer containing BSA (0.65 mg/mL of trypsin inhibitor final).
NOTE: Prepare a fresh trypsin inhibitor solution. - Prepare the ammonium persulfate reagent by adding 0.1 g of ammonium persulfate in 1 mL of water (10% ammonium persulfate final).
NOTE: The ammonium persulfate reagent should be either freshly prepared or a large volume can be aliquoted and kept in a -20 °C freezer. - Prepare the aminocaproic acid solution (ACA) by combining 0.131 g of 6-amino caproic acid in 1.0 mL of ultra-purified water and store it in 4 °C (1 M 6-aminocaproic acid final).
- Prepare the Coomassie brilliant blue dye solution by dissolving 1 g of Coomassie G-250 in 0.5 mL of water, and 0.5 mL of 1 M aminocaproic acid (10% dye solution final concentration).
- Prepare 5% Digitonin for Blue native-PAGE (BN-PAGE). Approximately calculate how much digitonin is needed in the sample preparation plus some extra volume to offset the pipetting errors (overestimate by 10 mg). For 15 mg: First, dilute 15 mg of digitonin in 30 µL of 100% DMSO. Vortex to help the digitonin to get into the solution. Next, add the remaining 90% of ddH2O (270 µL). Gently heat to mix the solution (vortex), and then cool it on ice.
- Prepare the native PAGE anode buffer. Add 50 mL of 20x native PAGE running buffer to 950 mL water to make a final volume of 1 L. Prepare fresh anode buffer for immediate use. Use this as a 1x running buffer.
- Prepare a dark blue cathode buffer. Dissolve 0.0160 g of Coomassie brilliant blue G-250 in 80 mL of native PAGE anode buffer; mix well.
NOTE: Prepare fresh cathode buffer for immediate use. Only 60 mL may be needed but make an extra 20 mL in the event of any leaks. - Prepare the light blue cathode buffer. Add 20 mL of dark blue cathode buffer into 180 mL of native PAGE anode buffer and mix well.
NOTE: This buffer can only be used once. - Prepare 5% Coomassie G-250 as a sample additive. Dilute 200 µL of 10% Coomassie stock already made (step 1.8) with 200 µL of 1 M ACA and store at the 4 °C.
- Prepare 4x native PAGE sample buffer by combining 40% glycerol, 0.04% Ponceau S, 25 mM Tris-HCl (pH 6.8), 192 mM glycine in 10 mL of final volume. Aliquot them and store at -20 °C in the freezer.
2. Mitochondria isolation from frozen heart tissue
NOTE: Perform the mitochondria isolation procedure in a 4 °C cold room. However, in case of unavailability of the cold room, use a large size ice bath to perform the procedure.
- Take out previously frozen heart tissue samples from the -80 °C freezer.
NOTE: A maximum of four tissue samples may be comfortably processed for mitochondria isolation in a given day. - Weigh the tube containing the heart tissue sample and record the weight. Prepare three beakers (each 50 mL) containing 40 mL of washing buffer. Cool each beaker in the ice-salt bath with equal amounts of ice and salt mixed for 3-5 min before proceeding to the next step. Put magnetic stirring bars in each beaker and set the stirrer for medium speed.
NOTE: Do not over-freeze the beakers and use them immediately just after clouds of ice crystals start to appear. - Put the frozen heart tissue directly into the ice-cold solution in the first beaker. Wait approximately 5 min while stirring and applying slight pressure to the tissue using tweezers to get out as much blood as possible.
- Weigh the empty tube and subtract from the original weight (step 2.2) to obtain a net weight of the heart tissue.
- Take the tissue out with forceps and squeeze the heart with a clean paper towel of filter paper to remove blood. Then, place the tissue into the second beaker. Wait approximately 5 min while stirring to remove excess blood.
- Gently dry the tissue on a paper towel. Remove all fat, clotted blood, auricles, and fasciae (white tissue). Then, pool the ventricular tissue.
- Shred the tissue samples using the shredder by following steps 2.7.1-2.7.9.
- Place the heart tissue (~50-300 mg) into the tissue shredder chamber from the ram side.
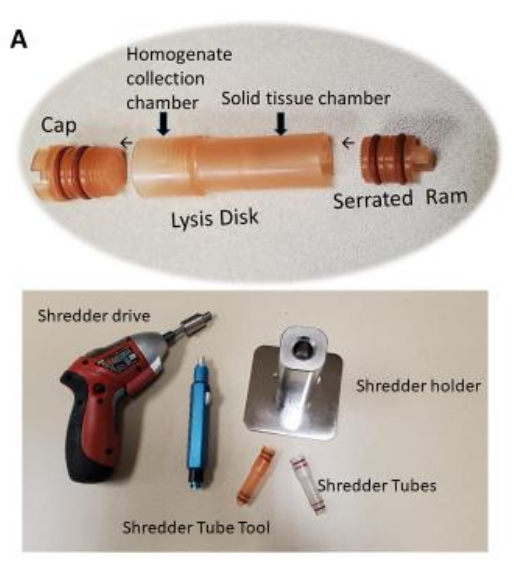
Figure 1: Tissue shredder chamber (tube) for use with the shredder. Tissue homogenization in the shredder tube requires about 10-12 s per tissue sample. Tissue homogenization is completed once the homogenized tissue passes through the perforated disk into the upper chamber. Please click here to view a larger version of this figure.
- Add ~0.2-0.8 mL of washing buffer to the cap side of the tube.
NOTE: Ensure that the total volume of the sample and the washing buffer combined during the shredding does not exceed 0.7-0.8 mL. - Use the tube tool to cap the shredder chamber with the shredder cap snugly.
NOTE: Do not over tighten the shredder cap. - Place the shredder chamber into the shredder holder unit with the ram side down. The ram will engage a fitting in the shredder holder.
- Insert the bit of the shredder driver; let the bit engage with the shredder cap. Make sure that the bit is snugly locked into place.
NOTE: Always use the fully charged shredder driver. - Manually apply pressure downward to the shredder driver and shredder chamber until the pressure indicator on the shredder holder reaches a setting of approximately 2 on the shredder holder.
NOTE: The setting can be at a higher level if shredding tissue is more fibrous. - Run the shredder driver for approximately 10-12 s by pressing the trigger of the shredder driver, while maintaining the setting at 2.
- Look at the tube to check whether all or most of the solid mass of the tissue is shredded through the lysis disk and passed into the upper chamber of the shredder chamber by looking at the tube.
NOTE: It may be necessary to shred the tissue sample for a longer period. Simply return the tube to the shredder holder and continue the process. Most kinds of tissue samples require only 10-12 s of shredding. While all shredding will produce some heat due to friction, this short processing time will not significantly heat the sample. - After shredding the sample, lift the shredder chamber out of the holder unit. Use the tube tool to remove the shredder cap and place it on a clean surface for later use.
NOTE: The shredder cap should be considered contaminated with sample extract, and depending on the sample type, may be contaminated with infectious materials. Do not use the same shredder cap for any other tissue samples to prevent cross-contamination.
- Transfer the shredded heart tissue into the third beaker with 40 mL of washing buffer and stir bar and stir the tissue for 5 min at a medium speed while on the stir plate.
- Filter the suspension through a nylon mesh monofilament (74 µm pore size) and discard the filtrate. Wash the shredded tissue on the filter 3x with 10 mL of washing buffer.
NOTE: Make sure that the nylon mesh monofilament is pre-wetted with water. - Transfer the washed and shredded tissue into a 50 mL beaker with 20 mL of washing buffer and place the beaker in the ice bath on the magnetic stirrer. Add 0.5 mL of trypsin solution while constantly stirring for 15 min. Approximately halfway through the incubation (at about 7.5 min), transfer the tissue suspension into a loosely fitted handheld glass-on-glass homogenizer that will hold 0-15 mL of volume.
Figure 1: Tissue shredder chamber (tube) for use with the shredder. Tissue homogenization in the shredder tube requires about 10-12 s per tissue sample. Tissue homogenization is completed once the homogenized tissue passes through the perforated disk into the upper chamber. Please click here to view a larger version of this figure.
- Gently homogenize with 4-5 strokes to disperse the suspension without producing froth. Place the homogenate into a 50 mL beaker and gently mix on a stirring plate for 15 min at 4 °C.
- After 15 min of incubation, add 10 mL of the isolation buffer that contains the trypsin inhibitor into the beaker containing the homogenate and incubate for 1 min at 4 °C while gently stirring.
- Filter the suspension again through a nylon filter and save the filtrate in a 50 mL conical tube.
- Collect the particulate fraction left on the nylon filter and place it in the glass-on-glass homogenizer. Add 15-20 mL of the washing buffer and gently homogenize by hand for 25-30 s.
NOTE: The filtrate flows very slowly at this step. To increase the filtrate flow, place the funnel tip touching the inside of the conical tube. - Combine the homogenate with the filtrate, balance the tubes, and centrifuge at 600 x g for 15 min at 4 °C.
- Filter the resulting supernatant into a new conical tube using a nylon filter to avoid contamination by the pellet. Run the centrifuge again at 8,500 x g for 15 min at 4 °C.
NOTE: Use a plastic transfer pipette to remove the supernatant and sacrifice 0.2 mL of supernatant from the top of the fluffy pellet. - Discard the supernatant and rinse the pellet 2-3 times with 1 mL of the isolation buffer. Ensure to discard the fluffy white outer rim layer each time.
NOTE: Wash the pellet by adding the isolation buffer using a pipette with 1 mL tip, or a new transfer pipette. Add the buffer to the side of the tube, not directly over the pellet, to prevent breaking the pellet and prevent the contamination of the supernatant. Next, aspirate the buffer with a transfer pipette and repeat the process two more times. - Add 5 mL of the isolation buffer to each tube and break up the pellet using a pipette with a 1 mL tip, or a new transfer pipette.
- Dilute the suspension with 20 mL of additional isolation buffer and run the centrifuge again at 8,500 x g for 15 min at 4 °C.
- Discard the supernatant and resuspend the pellet in 5 mL of the resuspending buffer. Then, add 15 mL more of the buffer for a total of 20 mL, and centrifuge at 8,500 x g for 15 min. at 4 °C. Discard the supernatant. The resulting brown pellet contains washed intact mitochondria.
- Resuspend the mitochondria-enriched pellet in 250 µL of the resuspending buffer, including protease inhibitors. Aliquot in 25 µL, quick freeze in liquid nitrogen, and store in a -80 °C deep freezer for several years.
NOTE: Spare 5 µL of mitochondria-enriched preparation for the protein analysis assay. For a BN-PAGE (Blue native-PAGE) study, aliquot the resuspended pellet in 50 µg of mitochondrial protein per tube. Centrifuge at 8,500 x g for 15 min at 4 °C. Carefully aspirate the supernatant and store the mitochondrial pellet (50 µg protein/aliquot) in a -80 °C deep freezer for several years.
3. Mitochondrial electron transfer complex (ETC) assessment
NOTE: Individual mitochondrial ETC proteins (i.e., Complex-I, II, III, IV, V) can be assessed by standard immunoblotting assay. Due to the variety of samples (four time points: 48 h, 3 months,6 months, 9 months), two experimental conditions (exercised and sedentary), and a number of immunoprobing antibodies (five antibodies), a multiplexed immunoblotting is recommended. Perform multiplexed immunoblotting as follows.
- Resolve 50 µg of mitochondria enriched protein sample/well in 4%-20% gradient polyacrylamide gel electrophoresis (100 V, 90 min).
- Transfer the proteins onto a PVDF membrane (75 V; 60 min)
- Block the membrane in a blocking buffer solution for a minimum of 1 h at 4 °C.
NOTE: Blocking time can be extended for up to overnight at 4 °C. The blocking buffer (Table of Materials) contains non-mammalian ingredients (i.e., fish proteins) that are less likely to have specific binding interactions with antibodies as well as other mammalian proteins present in typical methods. - Probe the membrane with the antibody cocktail including anti-Complex-I, II, III, IV, V antibodies and incubate overnight on an orbital shaker at 4 °C.
- Next day, wash the membrane and probe with IR-tagged secondary antibody for 2 h on an orbital shaker at RT (room temperature).
- Wash five times with 1x TBS-T (TBS containing Tween 20), and once with 1x TBS (Tris buffered saline).
NOTE: For the last wash, do not include detergent in the TBS buffer. - Image the blot in an Image analyzer.
4. The mitochondria supercomplex assessment by Blue Native Poly Acrylamide Gel Electrophoresis (BN-PAGE)
NOTE: A supercomplex profile of ETC can also be assessed by employing the BN-PAGE technique.
- Prepare the sample buffer cocktail for 20 µL per 50 µg mitochondria-enriched pellet protein (7 µL of water + 5 µL of 4x native polyacrylamide gel electrophoresis sample loading buffer + 8 µL of 5% Digitonin + 2 µL of Coomassie G-250).
NOTE: Make a stock sample buffer cocktail depending on the number of samples available (50 µg protein/sample). Choose appropriate digitonin amounts based on the protein concentration of the mitochondria-enriched pellet (Table 1)8. - Heat the digitonin stock at 95 °C for 3 min, mix, and cool on ice before making the master mix.
- Add the Coomassie G-250 sample additive just before loading the samples into the gel.
- Add 20 µL of the sample buffer cocktail in a tube that contains 50 µg of mitochondrial protein pellet. Solubilize/resuspend the pellet by gently mixing the pellet with a pipette without generating froth.
- Incubate the mixture on ice for 20 min to achieve maximum solubilization of the mitochondria-enriched pellet. From time to time, gently flick the tubes to enhance solubilization.
- While waiting for incubation, prepare the native PAGE anode buffer/1x running buffer and the dark blue colored cathode buffer.
- Centrifuge the solubilized mitochondria-enriched pellet at 20,000 x g for 10 min at 4 °C.
- After centrifugation, collect 15 µL of the supernatant into new tubes placed on ice.
- Add the appropriate amount of 5% Coomassie G-250 reagent to the supernatant obtained in step 4.8 (refer to Table 1).
NOTE: The 5% Coomassie G-250 volume should be such that the final concentration of the dye is ¼ the detergent concentration. Perform this step just before loading the samples into the gel.
| Protein | Digitonin/protein | 4x Sample Buffer | 5% Digitonin | Water | Final volume | 5% Coomassie G-250 |
| ratio (g/g) | sample additive | |||||
| 50 µg | 8 | 5 | 8 | 7 | 20 | 2 |
| 50 µg | 4 | 5 | 4 | 11 | 20 | 1 |
Table 1: Sample Buffer Cocktail Preparation with two different detergent/protein ratios. To achieve maximum solubilization of membrane proteins from the mitochondrial suspension, the digitonin/mitochondrial suspension protein concentration should be adjusted to between 4-8 g digitonin/g of protein. In the heart tissue, 400 µg digitonin for 50 µg of mitochondrial suspension proteins (i.e., 8 g digitonin/g of mitochondria protein) were used to maximize the solubility of the supercomplexes from the mitochondria suspension.
- Pour a 3%-8% gradient gel, remove the comb, and wash out the wells of the gel with 1x running buffer three times.
NOTE: The gel best polymerizes if poured a day before. - Set up the electrophoresis system and place the gel in the apparatus.
- Fill the sample wells with the dark blue colored cathode buffer.
- Load 10 µL of the unstained protein standard as a native molecular weight marker on the first and last well of the gel.
- Load the entire amount of the sample supernatant + Coomassie sample additive prepared in step 4.9 above into the wells using a flat-head gel-loading tip.
- Carefully fill the mini cell buffer dam with approximately 60 mL of dark blue colored cathode buffer without disturbing the samples in the wells, and then fill the buffer tank with the native PAGE 1x running buffer.
- Turn on the power supply and electrophorese the samples at 150 V for approximately 30 min.
- Remove the dark blue color cathode buffer from the buffer dam (inner chamber) with a transfer pipette or aspirate the sample wells. Fill the inner chamber with 150-200 mL of light blue colored cathode buffer and run the electrophoresis at 250 V for 60 min.
NOTE: The electrophoresis time may be determined by checking when the dye front migrates just 0.5 cm above the bottom of the gel cassette. - Remove the gel from the cassette and place it in a clean plastic container. Wash the gel with good quality distilled water for 15 min on a rocker/orbital shaker.
NOTE: To remove the gradient gel from the cassette, handle the cassette from the bottom, which is stronger, to minimize the chances of breakage. - Pour off water, immerse the gel in a sufficient amount of Coomassie GelCode Blue stain reagent, and incubate for 1 h on an orbital/rocker at room temperature.
NOTE: Mix the Blue stain reagent (Table of Materials) solution before use by gently swirling the bottle several times. - Decant the dye solution, add distilled water, rinse several times with water, and then put on the orbital/rocker for destaining.
NOTE: Insert a piece of sponge (~3 cm x 0.5 cm) into the gel destaining container and leave the gel for overnight destaining. The sponge adsorbs the dye particles without requiring multiple water changes. - Analyze the gel in an image analyzer.
5. Mitochondria supercomplex assessment by immunoblot blot analysis
- Perform a new set of BN-PAGE without staining the gel at the end of the electrophoresis.
- Follow the same procedure as in section 3 (mitochondrial electron transfer complex (ETC) assessment) to complete the immunoblotting.
Following the protocol, a good yield of mitochondria-enriched protein mixture from heart tissue was prepared. Approximately 15 mg/mL of mitochondria-enriched protein mixture was obtained from an average of 1.2 g frozen heart tissue harvested from the offspring of the sow. Observations indicated that less than 0.5 g of frozen heart tissue did not yield a sufficient amount of mitochondrial-enriched protein mixture to carry out a BN-PAGE assay. The amount of mitochondrial preparation was sufficient to perform (i) a standard immunoblot analysis for assessing the individual electron transfer complexes (ETC) (i.e., Complex-I, II, III, IV, and Complex-V), (ii) a mitochondrial supercomplex assessment by BN-PAGE and immunoblot analysis, and (iii) limited enzymatic assays for the select complex. The cocktail of antibodies raised against individual ETC protein complexes provides technical feasibility that each antibody will recognize its own target protein in one single immunoblot assay.
Mitochondria-enriched protein was prepared from the left ventricle of the heart, obtained from the offspring of the sow. The offspring were born to two groups of sows: (1) one group was exercised during pregnancy and (2) one group was sedentary. The heart tissues of both the groups were harvested at different time points after the birth (48 h, 3 months, 6 months, and 9 months). Whether maternal exercise has a positive impact on mitochondrial ETC of the offspring during gestation was the hypothesis that was tested in this study. Four heart tissues were comfortably processed on any given day due to the lengthy mitochondria-enriched isolation process.
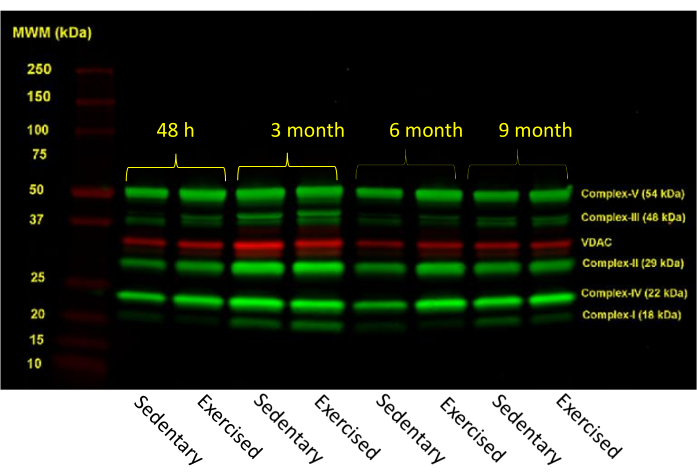
Figure 2: Immunoblot profile of mitochondrial ETC complexes from offspring heart tissue. The mitochondrial-enriched protein mixture was resolved in a 4%-20% gradient gel under reducing conditions. Proteins were immunoprobed by a commercial antibody cocktail, and anti-VDAC antibody was used for loading control (red color band). The cocktail of antibodies was raised against bovine heart Complex-I (antigen NDUFB8), bovine heart Complex-II (antigen C-II30 (FeS), bovine heart Complex II, III (antigen C-III-Core 2), human Complex IV, subunit II (antigen C-IV-II), and bovine heart Complex-V (antigen C-V-α). The secondary antibody was labeled with an infrared tag, and the image was analyzed using an image analyzing software. Individual Complex profiles across the age groups are provided in Supplemental Figure 1 (A–E). Please click here to view a larger version of this figure.
Figure 2 represents a typical ETC protein profile. Antibody cocktails allowed for immunoblotting in a multiplex format. Each antibody recognized its own target protein, providing an acceptable resolution. The protein band intensities were digitally analyzed. Tissues obtained from the offspring of exercised sow presented relatively reduced levels in some of the ETC complexes (Supplemental Figures 1A–E). The mitochondria-enriched preparation must be resolved in a gradient gel (4%-20%) as the immunoprobing was performed in a multiplexing format. This approach will eliminate the necessity of running several fixed percent of SDS/PAGE gels and run-to-run variability.
Mitochondrial proteins were resolved in a 3%-8% gradient BN-PAGE (Figure 3A) and immunoprobed with antibody cocktails. In both exercised and sedentary groups, multiple supercomplex formation was observed (Figure 3B).
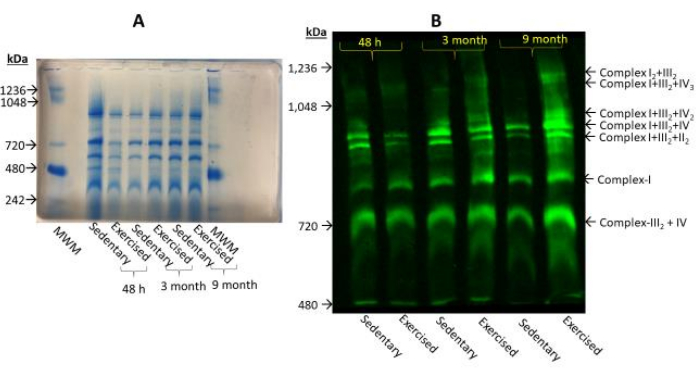
Figure 3: Mitochondrial supercomplex profile by Blue-Native Polyacrylamide gel electrophoresis (BN-PAGE). (A). Mitochondrial-enriched protein mixtures were resolved in 3%-8% BN-PAGE, and stained with a Coomassie blue reagent (Gel Code, Blue Stain Reagent). (B). Mitochondrial-enriched protein mixtures were resolved in a 3%-8% BN-PAGE, transferred onto a PVDF membrane, and immunoprobed with the antibody cocktails for supercomplexes. In both assays, the protein load was 150 µg/well. A 6-month data collection point was not included due to an insufficient protein yield during the preparation. The most prominent supercomplex increase observed was at 9 months in the exercised group compared to the sedentary group. Please click here to view a larger version of this figure.
A different gradient gel format (3%-8%) was prepared due to the large size of the supercomplexes that can be resolved in BN-PAGE. Mitochondria-enriched preparation described in this protocol was used for measuring mitochondrial ETC complex activities. Figure 4 represents data recorded for Complex I-II activity assessment. This assay was performed according to the established protocol3.
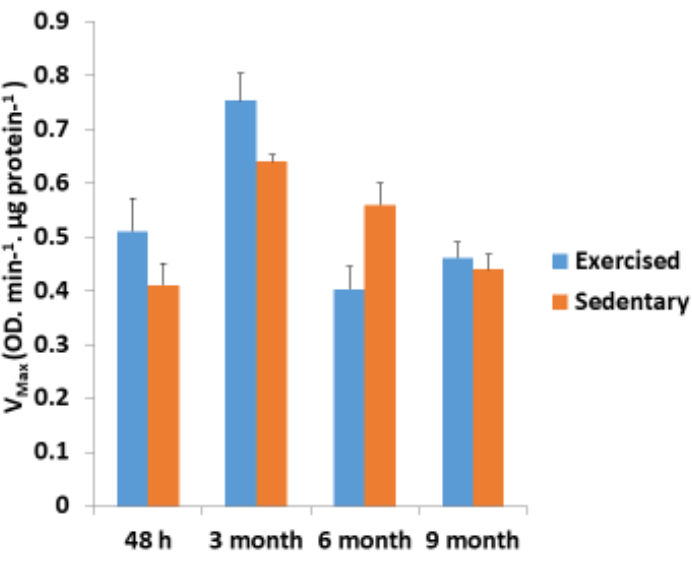
Figure 4: Complex I-II activity measurements. Mitochondria-enriched protein mixtures obtained from different age groups (48 h, 3 months, 6 months, and 9 months) were analyzed for the Complex I-II activity using a kinetic assay. The exercised group showed an increased Complex I-II activity as compare to that of the sedentary group. The difference between the two groups was not statistically significant according to a paired t-test (P > 0.05). The error bars depicted in the figure represent the standard error of the mean (SEM) for n = 4. Please click here to view a larger version of this figure.
Supplemental Figure 1: (A) Complex-I profile across age groups. In general, offspring born to the exercised sow presented with lower levels of Complex-I compared to that of the sedentary group (n = 4). This was more pronounced in the 9-month group. (B) Complex-II profile across age groups. All age groups showed low levels of Complex-II in the exercised group with the exception of three months cohort (n = 4). (C) Complex-III profile across age groups. Only the 9-month age group showed lower levels of Complex-III in the exercised group as compared to that of the sedentary group (n = 4). (D) Complex-IV profile across age groups. A slight increase in the Complex-IV protein levels of the exercised group was observed (n = 4). (E) The Complex-V (ATP synthase) profile across age groups. Both exercised and sedentary groups exhibited low levels of Complex-V except at the 48 h data point (n = 4). Please click here to download this File.
