A wild C. tropicalis strain (JU1848) was isolated from rotten palm tree fruits in the Nouragues Forest of French Guiana (Figure 1A)24. This strain was found to have thin microbes that colonize the lumen of the intestine in a directional manner (Figure 1B). This microbe was easily transferred to C. elegans strain N2 via co-culture with strain JU1848, where it colonized the lumen of the intestine similarly.
Propagation of JU1848 on standard NGM plates seeded with E. coli OP50-1 over multiple generations continually resulted in visible contamination, seen as various dark, mucoid colonies on and off the OP50-1 lawn (Figure 2A). A plate of wild JU1848 was starved to force animals into dauer and cleaned as described. Single dauer animals that survived cleaning were plated onto individual 6 cm NGM plates seeded with OP50-1 and allowed to grow for 4 days at 20 °C. Multiple plates of F1 progeny were observed without visible microbial contamination (Figure 2B). The F1 progeny were verified to still contain adhering bacteria in the lumen of the intestine (see below).
Clean JU1848 animals were washed and decapitated to isolate intestinal pieces as described in the protocol (steps 6.1-6.12). Adhering bacteria in the lumen of the dissected intestine was verified via Nomarski microscopy (Figure 3). The microbe in the lumen of JU1848 was suspected to be a bacterium, so the dissected intestines were used as a template for PCR using universal bacterial 16S primers, 27F, and 1492R. From a total of six individual dissected intestines, the PCR products were sequenced via Sanger, and clean chromatographs showed that all the six sequences were identical. Based on these sequences, this bacterium was identified as a new species in the class Alphaproteobacteria but could not be classified into a known order or genus (Supplementary File 1).

Figure 1: Adhering bacteria colonizing the lumen of a wild C. tropicalis. (A) Field sample image of rotten Euterpe sp. (Family: Arecaceae) palm tree fruits in the Nouragues forest of French Guiana. (B) Nomarski image of strain JU1848 seen with thousands of long, thin bacteria that form a brush-like appearance in the lumen (lu) of the host intestine (in). The bacterial (ba) layers coating the intestine are indicated with brackets ([). Please click here to view a larger version of this figure.
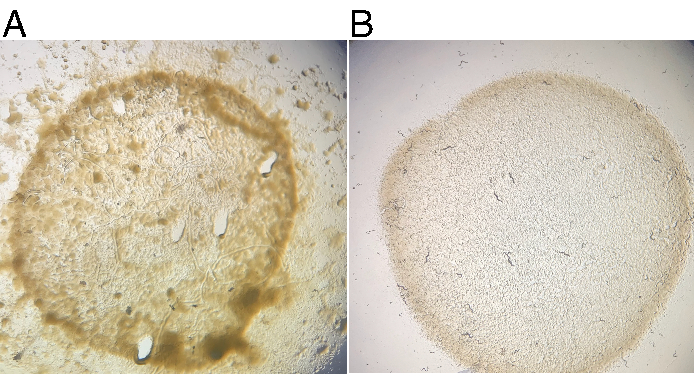
Figure 2: Contaminating microbial growth is lost after nematode cleaning. (A) Wild strain JU1848 propagates noticeable microbial growth on standard 6 cm NGM plates seeded with E. coli OP50-1 bacteria. (B) After cleaning, a plate of F1 progeny from a single dauer shows no visible microbial contamination after 4 days of incubation at 20 °C. Please click here to view a larger version of this figure.
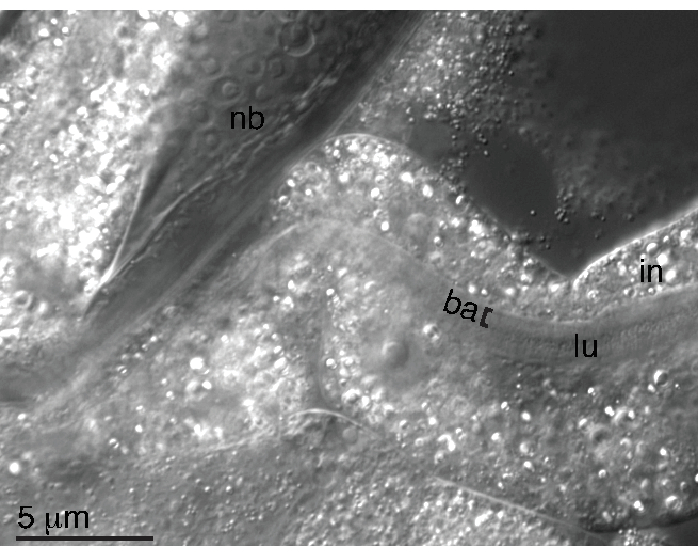
Figure 3: Adhering bacteria are seen in the lumen of the dissected intestine. Nomarski image of a clean JU1848 animal that was decapitated so that the intestines spill out. The colonizing bacteria (ba) are indicated with a bracket ([) and are seen in the lumen (lu) of a piece of the intestine (in) that is outside of the nematode body (nb). Please click here to view a larger version of this figure.
| Reagent | Concentration | Amount |
| 27F primer (5’-AGAGTTTGATCMTGGCTCAG-3’) | 20 mM | 2.5 µL |
| 1492R primer (5’-GGTTACCTTGTTACGACTT-3’) | 20 mM | 2.5 µL |
| Dissected intestine in water | N/A | 3 µL |
| 10x PCR buffer | 10x | 5 µL |
| dNTP | 10 mM | 1 µL |
| Taq Polymerase | 5 U/µL | 0.5 µL |
| Water | N/A | 35.5 µL |
Table 1: Sample PCR protocol using universal bacterial primers and dissected intestine.
Supplementary File 1: MUSCLE alignment of bacterial 16S rDNA sequences derived from PCR of six dissected JU1848 intestines. Please click here to download this File.
