Stab wound injury in the optic tectum using needle insertion into the right hemisphere (Figure 1, Figure 4A, and Figure 5A) induces various cellular responses, including radial glial cell (RGC) proliferation and the generation of newborn neurons. Similarly, aged populations of zebrafish and medaka were used to counteract any aging effects in the regenerative response. Then fluorescent immunostaining was performed on the frozen sections, and the RGC proliferation and differentiation were analyzed after the tectum injury in the zebrafish and medaka (Figure 4-5)13.
Antibodies against a proliferating cell marker were used, proliferating cell antigen (PCNA), and an RGC marker, brain lipid-binding protein (BLBP), available in zebrafish and medaka to evaluate the RGC proliferation in these tissues13,19. As previously described, most of the RGCs were quiescent (PCNA negative) in the contralateral uninjured hemisphere (Figure 4B)13. Still, RGC proliferation was induced at 2 days post-injury (dpi) in the medaka tectum (Figure 4C,D)13. Induction of RGC proliferation after the injury is a common feature of both the zebrafish and medaka regenerative responses13,19.
BrdU labeling is a simple method used to evaluate cell lineage and analyze RGC differentiation after brain injury (Figure 5A)13. Immunostaining with antibodies for a pan-neuronal marker, HuC, and BrdU was previously used to compare RGC differentiation in the injured tectum of zebrafish and medaka (Figure 5B,C)13. If these antibodies are available in the target species, comparative analyses can be performed.
If the injury is appropriately induced, the injury site is located in the central-dorsal region in the optic tectum (Figure 4C). Nuclear staining and hematoxylin and eosin staining can then be used to confirm the injury site13,19,20,21,22,23,24,25. After the injury, a disturbed periventricular gray zone with nuclear staining can be observed (Figure 4C). If the injury is located in the medial dorsal region, RGC proliferation is not significantly increased25.

Figure 1: Stab wound injury in adult optic tectum using a needle. (A) Dorsal view of an adult zebrafish. Zebrafish is kept upright between two bent needles inserted vertically into a Styrofoam. (A') Magnified image of the boxed area in (A). 30 G needle is inserted into the medial region of the border between two skulls called os frontale and os parietale on the optic tectum. The yellow circle indicates the injury site, and white dashed lines indicate the two skulls on the optic tectum. (B) Dorsal view of adult medaka. (B') A magnified image of the boxed area in (B). 30 G needle is inserted into the border on the optic tectum. The yellow circle indicates the injury site, and white dashed lines indicate two skulls on the optic tectum. (C) Medaka has scales on the skull. Skull scales are to be removed before the stab wound injury. The dashed line indicates the scale on the optic tectum. Scale bar: 2 mm in A-B, 1 mm in A', B' and C. Telencephalon (Tel), optic tectum (OT), os frontale (F), and os parietale (P). Please click here to view a larger version of this figure.
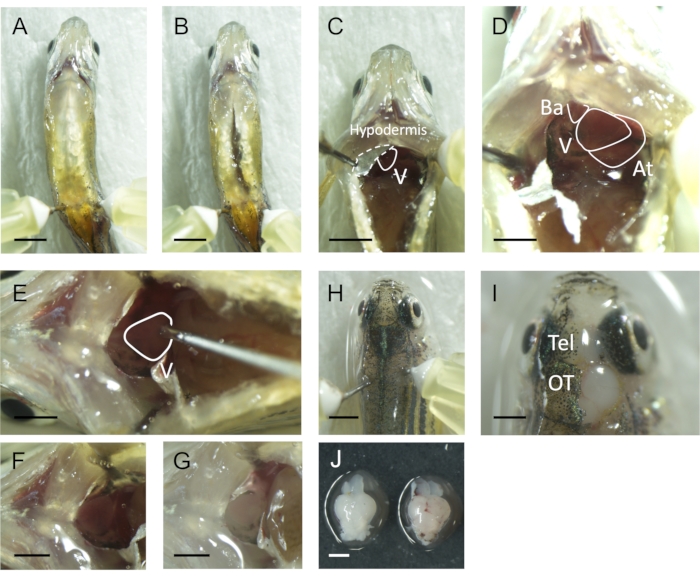
Figure 2: Intracardiac perfusion in small adult fish. (A) Ventral view of a zebrafish fixed on Styrofoam using bent needles ready for intracardiac perfusion and brain dissection. (B) A ventral incision is made from the origin of the anal fins to the chest. (C) The heart is behind a silvery epithelial layer called the hypodermis in both zebrafish and medaka. Another fixation using a bent needle beside the silver epithelial layer allows for easier access. The solid white line indicates the ventricle (V), and the dotted line indicates the hypodermis. (D) The silver epithelial layer is removed before the intracardial perfusion of 1x PBS. (E-G) Canula is inserted into the ventricle for intracardiac perfusion. Gills before (F) and after (G) the intracardiac perfusion. If the blood removal is not complete, the gills remain red. (H-J) Brain dissection after PBS perfusion to remove the blood from the tissues. Remove skulls on the optic tectum and telencephalon as shown in (I). If the blood removal is not complete, the brain looks light pink (the right brain in (J)). Scale bar: 2 mm in A-C and H, 1 mm in D-G and I-J. The olfactory bulb (OB), telencephalon (Tel), optic tectum (OT), bulbus arteriosus (Ba), ventricle (V), and atrium (At). Please click here to view a larger version of this figure.
Figure 3: Brain embedding for frozen sections. (A) The brain is embedded in a cryomold with an embedding compound. Anterior is down. (B) Cryomolds are cooled on a precooled aluminum block. Telencephalon (Tel), optic tectum (OT). Please click here to view a larger version of this figure.
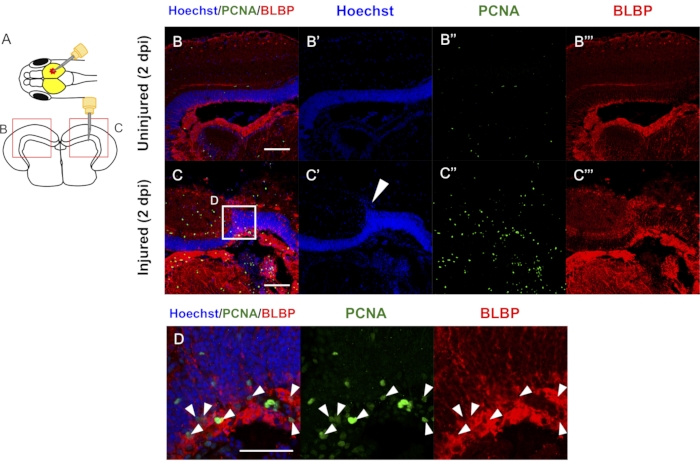
Figure 4: Representative results of fluorescent immunostaining against RGC proliferation after tectum injury in adult medaka. (A) Schematic view of the stab wound injury to the right hemisphere of the optic tectum and coronal section. (B-C) Representative results of proliferative RGCs (PCNA + BLBP + cells) in the contralateral uninjured (B) and injured (C) side at 2 days post-injury. White arrowhead in C' indicates a disturbed periventricular gray zone by the stab wound injury. (D) Magnified images of the boxed area in (C). White arrowheads in (D) indicate PCNA + BLBP + cells. Scale bar: 50 µm in B-D. Adapted with permission from Reference13. Please click here to view a larger version of this figure.
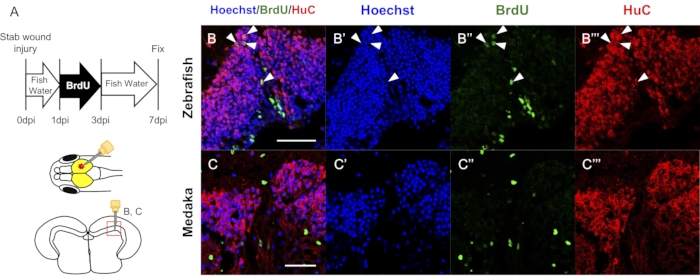
Figure 5: Representative results of the fluorescent immunostaining for the generation of newborn neurons after tectum injury. (A) Schematic view of bromodeoxyuridine (BrdU) treatment and the stab wound injury in the optic tectum and coronal section. (B-C) Representative results of newborn neurons (BrdU + HuC + cells) at 7 days post-injury in the injured zebrafish (B) and medaka (C). Scale bar: 50 µm in B-C. Adapted with permission from Reference13. Please click here to view a larger version of this figure.
