1. Preparation of clearing solutions
- To prepare CS solution, dissolve 10% (w/v) xylitol, 15% (w/v) sodium deoxycholate, and 25% (w/v) urea in distilled water on a magnetic stirrer.
NOTE: Sodium deoxycholate powder should be weighed in a draft chamber as it easily floats in the air. CS can be stored at room temperature in dark for more than 1 year. - To prepare CSA solution, add sodium sulfite (50 mM final concentration) to the CS solution obtained above.
NOTE: Add sodium sulfite to the CS solution right before use as the reducing agent gets deactivated easily.
2. Preparation of the fixative solution
- Transfer 40 mL of sterilized water into a conical tube and add 2 g of paraformaldehyde. Add 200 µL of 2 N NaOH to increase the pH of the solution. After closing and sealing the tube with parafilm, incubate at 60 °C with occasional inversions until everything is dissolved.
- After cooling the solution to room temperature, add 5 mL of 10x phosphate-buffered saline (PBS) to adjust the pH to 7.4. Add sterilized water to make up the volume to 50 mL.
NOTE: A freshly prepared fixative solution is preferred. The solution can also be stored at -30 °C for several months.
3. Fixation of samples
- Immerse plant samples in the fixative solution in a microtube (Figure 1A), ensuring that the volume of the fixative solution is more than five times the sample volume.
- Seal the microtube with a parafilm and make holes using a needle. Do not leave the tube open owing to a risk of sample spillage during vacuum decompression.
- Place the microtube in a desiccator and slowly adjust the degree of vacuum (~690 mmHg) so that bubbles appear gradually from the samples (Figure 1B-C). Turn off the vacuum pump after evacuating the desiccator. Leave the microtube undisturbed for 30 min at room temperature.
- Vent the desiccator carefully to prevent disturbing the samples. Turn on the vacuum pump again and turn it off after evacuating the desiccator. Leave the microtube undisturbed for 30 min at room temperature.
NOTE: Care should be taken to prevent damage to the samples while venting the desiccator. The penetration of the fixative solution into the samples is enhanced by two vacuum treatments. Further vacuum treatment helps to penetrate the fixative solution into thicker samples. - Open the desiccator carefully without bumping the fixative solution in the microtube. Using a micropipette, remove the fixative solution and add 1x PBS. After storing for 1 min, replace the old PBS with new 1x PBS (Figure 1D).
4. Clearing
- After removing PBS, add five times the sample volume of the clearing solution.
- Seal the microtube with parafilm and make holes using a needle. Place the samples in the desiccator, evacuate as in step 3.3, and turn off the vacuum pump. Leave the microtube undisturbed for 60 min at room temperature.
- Open the desiccator gently. Close the microtube with parafilm and store it at room temperature in the dark to avoid photobleaching of fluorescent proteins. Invert the microtube every 1-2 days to accelerate the clearing process.
- When the clearing solution turns green, replace it with new clearing solutions until the solution remains colorless (Figure 1E-H).

Figure 1: Procedure for CS treatment. (A) Arabidopsis seedling in 4% PFA (Paraformaldehyde) solution. (B) The sample is placed into a desiccator. (C) The seedling is fixed under a vacuum. (D) The seedling is soaked in the PFA solution after vacuum treatment. (E) Resulting 3-day clearing solution-treated seedling. Note the green color of the clearing solution. (F) The clearing solution is replaced 3 days after treatment. (G) Resulting 5-day clearing solution-treated seedling. (H) The clearing solution is replaced 5 days after treatment. Scale bars: 1 cm (A,C–H) and 5 cm (B). Please click here to view a larger version of this figure.
5. Chemical dye staining
- To the microtube, add Hoechst 33342 (final concentration of 10 µg/mL) for nuclear staining or Calcofluor White (final concentration of 1 mg/mL) for cell wall staining and wait for 1 h. After removing the dye solution, wash the sample with a fresh clearing solution for 1 h.
NOTE: Overnight staining and washing can improve fluorescent dye penetration into tissues and reduce background fluorescence. Various fluorescent dyes are compatible with the CS solution such as Basic Fuchsin9 (lignin), Auramine O9 (lignin, suberin, and cutin), Nile Red9 (suberin), Direct Yellow 969, Direct Red 239, and SR220010,11 (cell walls).
6. Observation
- Cut silicone rubber sheet with a razor blade to prepare a frame for the spacer (Figure 2A).
NOTE: Adjust the thickness of the silicon rubber sheet according to the thickness of the sample. As samples treated with clearing solution are soft, they will be damaged if they are covered directly with the cover glass. - Place the silicone frame on cover glass (e.g., 25 x 60 mm) (Figure 2B). Place the treated samples within the frame and add ~100 µL of clearing solution to remove any bubbles in the frame. Cover with another cover glass (18 x 18 mm or 24 x 24 mm) to prevent evaporation of the clearing solution (Figure 2C).
- Observe the samples under a fluorescent microscope. After observation, return the samples to the clearing solution taken in a microtube and store at room temperature in the dark.

Figure 2: Sample preparation for microscopic observation. (A) Cut a 0.2 mm thick silicone sheet into a frame. (B) Put the silicone sheet frame onto the cover glass. (C) Place the sample treated with clearing solution (marked by dotted border) within the frame and cover it with a cover glass. Scale bars: 5 mm. Please click here to view a larger version of this figure.
CS can clear leaves of various species (Figure 3A–H). It is difficult for the CS solution to penetrate a rice leaf because the leaf surface is covered by cuticular wax in this plant. However, after extracting the cuticular wax by dipping in chloroform for 10 s, CS could clear the rice leaves (Figure 3H). CS could not, however, penetrate Chamaecyparis obtusa leaves which are less permeable to CS (Figure 3I,J). In Chrysanthemum leaves, brown pigmentation induced by polyphenol oxidation was observed in the CS-treated leaves (Figure 3K,L). Similarly, Tobacco and torenia pistils showed brown pigmentation during the CS treatment (Figure 3M,O). As the sodium sulfite component in CSA prevents polyphenol oxidation owing to the reducing effect, CSA could clear tobacco and torenia pistils without any brown pigmentation (Figure 3N,P).
Figures 4B shows that CS treatment reduced the pale green color of the Arabidopsis H2B-mClover leaf (bright field) and enhanced the fluorescence intensity of H2B-mClover compared with PBS incubation (Figure 4A). In addition to flowering plants, CS is also applicable to moss plants (Figure 4C); after 4 days of CS treatment, the fluorescence of H2B-mRFP was clearly detected for the entire gametophore with reduced chlorophyll autofluorescence. Figures 4D,E show 3D reconstruction images of the H2B-mClover pistil in Nicotiana benthamiana cleared using CSA. The sample depth was 440 µm. As the depth-coded maximum intensity projection image shows, CSA allows for deep imaging of challenging tissues, such as tobacco pistils.
CS and CSA were also compatible with fluorescent dye staining. Figure 5 shows that CS could simultaneously be used to observe the fluorescent protein (H2B-mClover) and organic fluorescent dye staining (Calcofluor White). After 3D reconstruction from the z-stack images, any section could be observed.

Figure 3: Optical clearing of leaves and pistils using clearing solutions. (A–L) Fixed leaves of various species were incubated in PBS (A,C,E,G,I,K) or CS (B,D,F,H,J,L) for 8 days and CS (M,O) or CSA (N,P) for 2 days. (A,B,O,P) Torenia fournieri, (C,D) Nicotiana tabacum, (E,F) Cucumis sativus, (G,H) Oryza sativa, (I,J) Chamaecyparis obtusa, (K,L) Chrysanthemum morifolium, (M,N) Nicotiana benthamiana. Scale bars: 1 cm. Please click here to view a larger version of this figure.
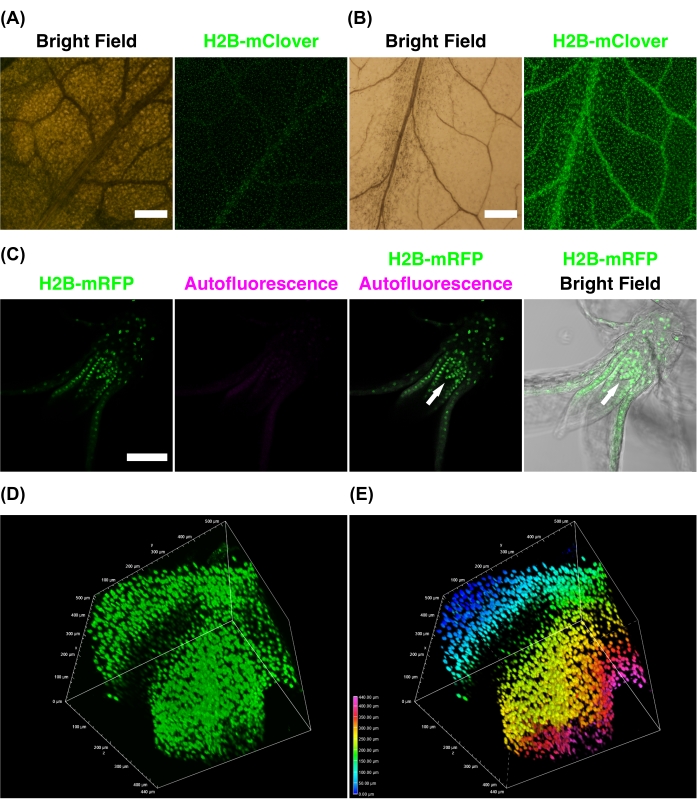
Figure 4: Fluorescence imaging of tissues treated with clearing solutions. (A,B) UBQ10pro::H2B-mClover leaves of Arabidopsis thaliana were treated with PBS (A) or CS (B) for 3 days. (C) H2B-mRFP leafy gametophores of Physcomitrium patens were treated with CS for 4 days. The nuclei were labeled with H2B-mRFP (green). The CS treatment reduced chlorophyll autofluorescence (magenta). The H2B-mRFP signal in the apical region was clearly observed for both merged images of H2B-mRFP and autofluorescence or bright field. (D,E) The UBQ10pro::H2B-mClover stigma of Nicotiana benthamiana was treated in CSA for 1 month. Maximum-intensity projection (D) and depth-coded maximum-intensity projection (E) were generated from 88 z-stack images at 5 µm intervals. Scale bars: 100 µm. Images were taken using wide-field (A,B), confocal (C), and two-photon excitation (D,E) microscopy. Please click here to view a larger version of this figure.
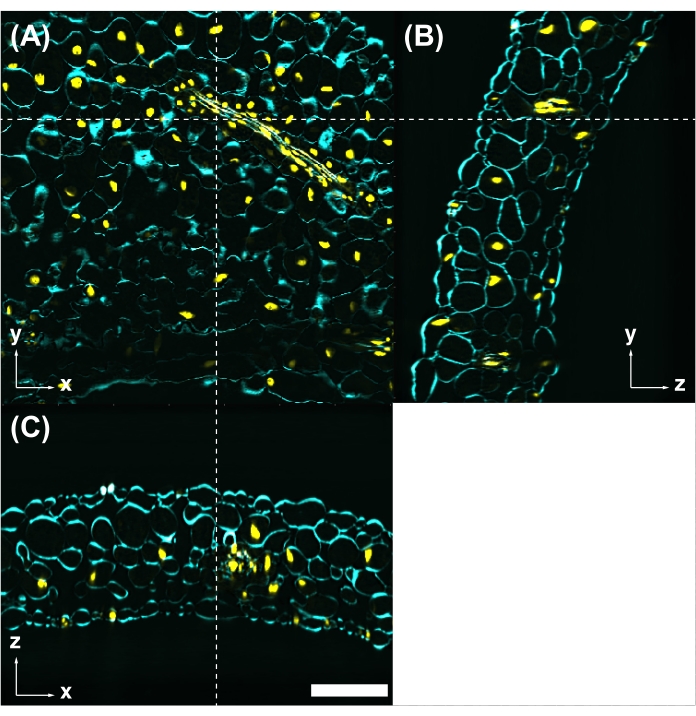
Figure 5: Fluorescent dye staining is compatible with CS. (A) CS-treated leaves observed by two-photon excitation microscopy with 950 nm excitation. Cell wall is stained with Calcofluor White (cyan). Nuclei are labeled with UBQ10pro::H2B-mClover (yellow). The yz (B) and xz (C) images are cross-sections at the position indicated by the white dashed lines in (A). Scale bar: 100 µm. Please click here to view a larger version of this figure.
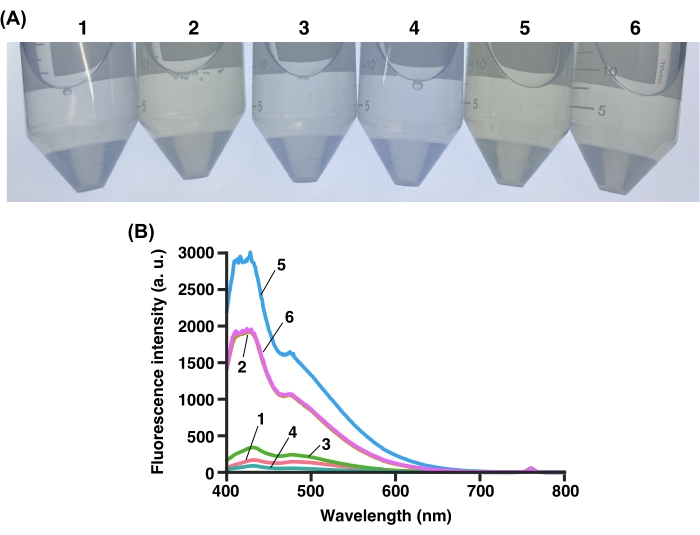
Figure 6: Transparency of sodium deoxycholate. (A) Colors of various 15% sodium deoxycholates (listed in Table of Materials). (B) Fluorescence spectrum of each 15% sodium deoxycholate with 380 nm excitation. Please click here to view a larger version of this figure.
