Typical O2 consumption and ΔΨ curves generated in these experiments are shown (Figure 1A,B). The logarithmic decline in the voltage signal with TPP+ calibration is shown at the start of each experiment. The absence of this logarithmic pattern may suggest a problem with the TPP+ selective electrode. Mitochondria typically generate ΔΨ immediately upon addition to respiratory buffer. ΔΨ can be interpreted from changes in TPP+ voltage based on the Nernst equation (logarithmic ratio of [TPP+ inside mitochondrial matrix] to [external TPP+])30. Note that physiological ΔΨ approximates the 2 μM TPP+ level of the standard curve. Voltage thresholds were thus defined relative to the 2 μM TPP+ standard (denoted as Δ mV TPP+ voltage signal).
For example, "high ΔΨ" was defined as ~Δ0 mV (at the 2 μM TPP+ level), an "intermediate ΔΨ" was set at ~Δ5 mV (below 2 μM TPP+), and "low ΔΨ" was set at ~Δ10 mV (below 2 μM TPP+) (Figure 1A,B). In pilot experiments, mitochondria at Δ0 mV exhibited 100% mPTP closed probability and those at Δ10 mV exhibited 100% open probability; representative graphs are shown in Figure 1A,B. The Δ5 mV threshold was thus arbitrarily chosen as an intermediate ΔΨ. Note that in these experiments, upon addition of mitochondria to the reaction chamber, mitochondria are in state 2 respiration where they are exposed to excess substrate. Mitochondria are not stimulated to enter state 3 with ADP; as a result, once oligomycin is added, they transition directly to state 4 oligomycin (leak respiration). As a result, an appreciable difference in oxygen consumption is typically not observed after oligomycin addition, suggesting that the ATP synthase contributes minimally to respiration during state 2 (Figure 1A,B). To evaluate the open or closed status of the mPTP, the O2 consumption rate and membrane potential just before and just after CsA addition are compared (Figure 1C). A decrease in the O2 consumption rate and an increase and stabilization of ΔΨ in response to CsA indicate closure of an open mPTP (blockade of pore-mediated proton leak) (Figure 1C, black curves).
When the mPTP is closed, there is no decrease in O2 consumption rate, and ΔΨ does not increase and stabilize but continues to fall (Figure 1C, red curves). The experimental results demonstrate similar closed and open mPTP probabilities at high and low ΔΨs, respectively, in FVB control and Fmr1 KO cardiac mitochondria (Figure 1D). These findings are consistent with known voltage-sensitive properties of the mPTP, where the pore tends to be closed at high ΔΨs and open at low ΔΨs13. Thus, using this method, we reliably demonstrated the usual voltage dependence of the mPTP in both strains. Interestingly, at the intermediate ΔΨ (Δ5 mV threshold), Fmr1 KO cardiac mitochondria demonstrated increased closed mPTP probability compared to FVB controls (Figure 1D). Thus, Fmr1 KO cardiac mitochondria demonstrated a shift in gating such that the pore required a lower ΔΨ for opening to be triggered. This is consistent with the previous finding that Fmr1 KOs have elevated levels of CoQ and a relatively closed mPTP26. Thus, the method identified an optimal ΔΨ threshold to resolve differences in mPTP opening. Importantly, defining this ΔΨ threshold will enable further investigation to better understand how CoQ regulates the mPTP and how CoQ deficiency leads to pathological pore opening in FXS.
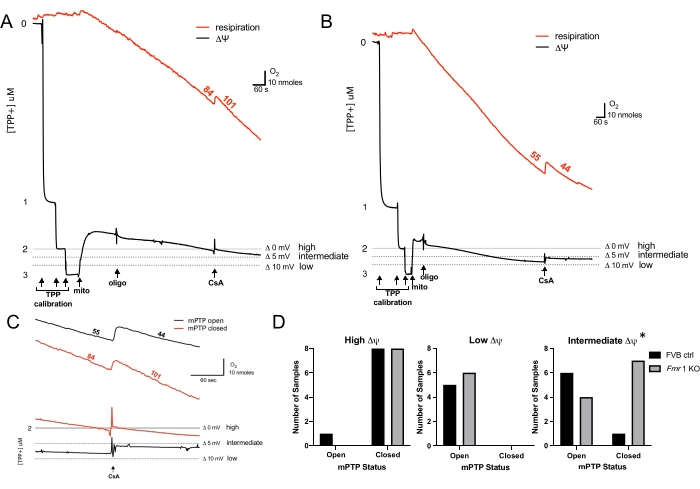
Figure 1: Detection of low-conductance mPTP open probability. (A, B) Representative curves of simultaneous O2 consumption (red, numbers are rates in nmol of O2 mL-1 min-1 mg protein-1) with ΔΨ (black, [TPP+]). Arrows indicate the addition of TPP+, mitochondria, oligomycin, and CsA. High, intermediate, and low voltage thresholds relative to the 2 μM TPP+ calibration level are shown. Addition of CsA at high (A) and low (B) voltage thresholds is shown. (C) Magnified section of O2 consumption (upper) and ΔΨ (lower) curves in (A) and (B) at the point of CsA addition, illustrating closed mPTP (red) at high ΔΨ (insensitive to CsA) and open mPTP (black) at low ΔΨ (sensitive to CsA). (D) Summary graphs of mPTP open and closed status at high, low, and intermediate ΔΨs are shown for FVB controls (black) and Fmr1 KOs (gray). Five to 11 animals were evaluated at each ΔΨ threshold; p values were calculated using a chi-squared test, *p < 0.05. Abbreviations: ΔΨ = mitochondrial membrane potential; mPTP = mitochondrial permeability transition pore; TPP+ = tetraphenylphosphonium; mito = mitochondria; oligo = oligomycin; CsA = cyclosporine A; ΔΨ = ; KO = knockout. Please click here to view a larger version of this figure.
| Component | MI/BSA* | SB/BSA* | Density Gradient Diluent | RB/BSA* |
| Mannitol | 225 mM | – | – | – |
| Sucrose | 75 mM | 250 mM | 1 M | 200 mM |
| HEPES | 5 mM | 5 mM | 50 mM | 5 mM |
| EGTA | 1 mM | 0.1 mM | 10 mM | – |
| KCl | – | – | – | 25 mM |
| KH2PO4 | – | – | – | 2 mM |
| MgCl2 | – | – | – | 5 mM |
| pH** | 7.4 | 7.4 | – | 7.2 |
| BSA*** | 0.10% | 0.10% | – | 0.02% |
| AP5A*** | – | – | – | 30 mM |
Table 1: Buffers and solutions.
* Filter through 0.2 μm filters
**Adjust pH with 3 M potassium hydroxide as needed.
***Denotes buffer components that are added just prior to mitochondrial isolation.
Abbreviations: MI = mitochondrial isolation buffer; SB = sucrose buffer; RB = Respiration buffer; BSA = fatty acid-free bovine serum albumin; HEPES = 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid; EGTA = ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid; KCl = potassium chloride; KH2PO4 = potassium dihydrogen phosphate; MgCl2 = magnesium chloride; AP5A = P1,P5-diadenosine-5' pentaphosphate pentasodium.
