The success of isolation of uMSCs from cord tissue is >95%, unlike the poor rates of success from whole cord blood. Upon successful isolation of uMSCs, FACS analysis reveals that all the cells are CD34−CD45−CD105+CD90+. However, in comparative analysis, uMSCs isolated from cord blood display heterogeneous populations, wherein a proportion of cells show CD34+CD45+CD105+ (~15%). Additionally, double-positive CD105+CD90+ are fewer in number (~5%) (Supplemental Figure S1). This results in reduced levels of myogenic differentiation amongst uMSCs from cord blood, as both CD105 and CD90 expression are required for the induction of myogenesis. uMSCs also display the expression of the markers listed in Table 1. If there is contamination of the cord blood-derived uMSCs, there will also be a presence of CD34+CD45+ cells. This results in false cell counts for plating for myogenic differentiation. An indication that >90% of the cells are double positive for CD105 and CD90 expression is that the uMSCs have not committed to any mesodermal lineage and continue to remain multipotent, as both CD105 and CD90 expression are downregulated upon differentiation. It is imperative to use multiple markers for the confirmation of uMSC phenotypes, as there is no single definitive uMSC marker. In this analysis, we evaluated the presence of all the markers listed in Table 1 for each of the uMSC lines established in the laboratory. Additionally, we determined the double-positive status of CD105 and CD90 (Figure 2A), as well as of CD105 and CD73 (Figure 2B), ensuring that the uMSCs express multiple key markers. This is necessary to avoid contaminating single-positive cells that are present in numbers of less than 0.05%.
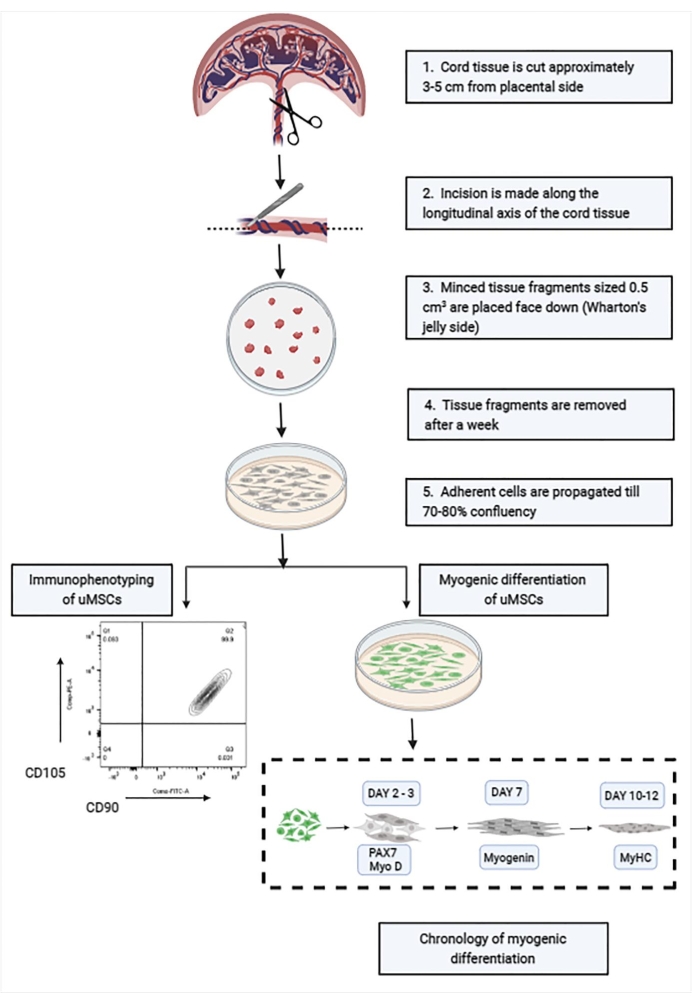
Figure 1: Schematic showing the stepwise isolation and characterization of uMSCs from cord tissue. Abbreviation: uMSCs = mesenchymal stem cells from human umbilical cord tissue. Please click here to view a larger version of this figure.
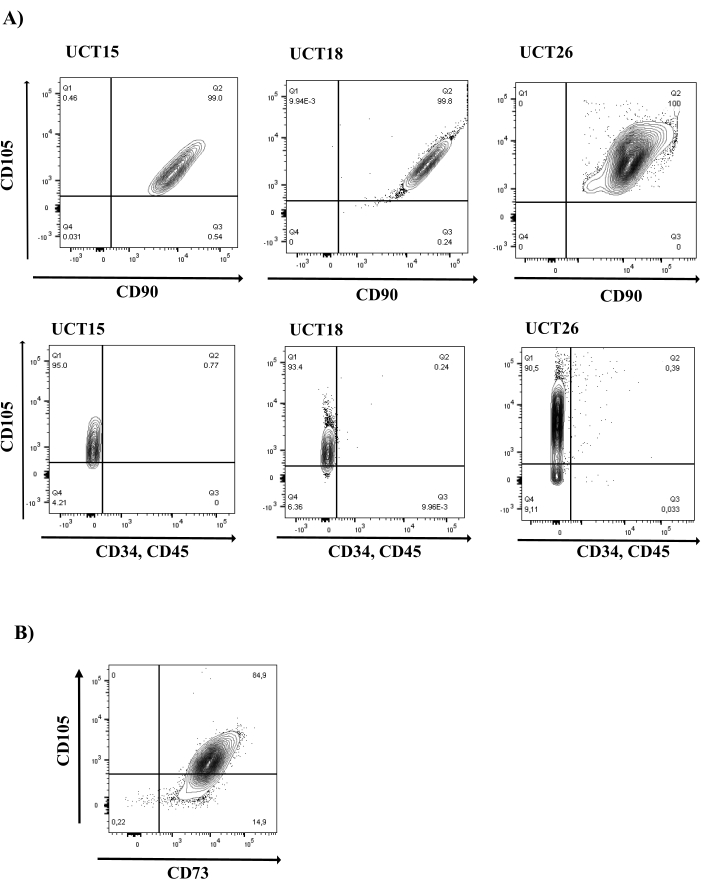
Figure 2: MSC marker analysis in cord tissue-derived uMSCs. (A) uMSCs display the expression of CD105 and CD90 and do not express hematopoietic markers, CD34 and CD45. Representative FACS plots of three uMSC lines (UCT15, UCT18, and UCT26) are shown (N = 16). Top row of panels shows cells in all three uMSC lines in the Q2 quadrant positive for CD105 and CD90 expression. Bottom row of panels shows cells in the Q1 quadrant positive for CD105 expression and negative for CD34 and CD45 expression. (B) uMSCs display the expression of MSC marker, CD73 (N = 16). Abbreviation: uMSCs = mesenchymal stem cells from human umbilical cord tissue. Please click here to view a larger version of this figure.
| uMSC positive markers | uMSC negative markers |
| CD105 | CD34 |
| CD90 | CD45 |
| CD73 | CD106 |
| CD29 | HLA DR |
| CD44 | CD31 |
| HLA ABC | CD14 |
| CD49e | CD49e |
| CD54 | |
| CD13 |
Table 1: List of positive and negative markers for uMSCs. Abbreviation: uMSCs = mesenchymal stem cells from human umbilical cord tissue.
For the differentiation of uMSCs into the myogenic lineage, uMSCs typically express Pax7, a marker for precursor cells within the first 2 days of the addition of M1, followed by MyoD within the first 4 days of M1 addition (Figure 3). At 6 days of differentiation, cells express Myogenin protein, followed by the expression of Myosin heavy chain (MyHC) between 10 days and 14 days of the induction of differentiation. We characterized in more detail the kinetics of myogenic expression using RNA sequencing, flow cytometry, immunocytochemistry, RT-PCR, and western blot analysis to document the stagewise expression of myogenic markers, confirming the robustness of this protocol. Whole-genome transcriptomic sequencing between undifferentiated uMSCs and uMSCs that were differentiated into skeletal muscle revealed the upregulation of 907 genes in response to the induction of myogenic differentiation (Figure 4).
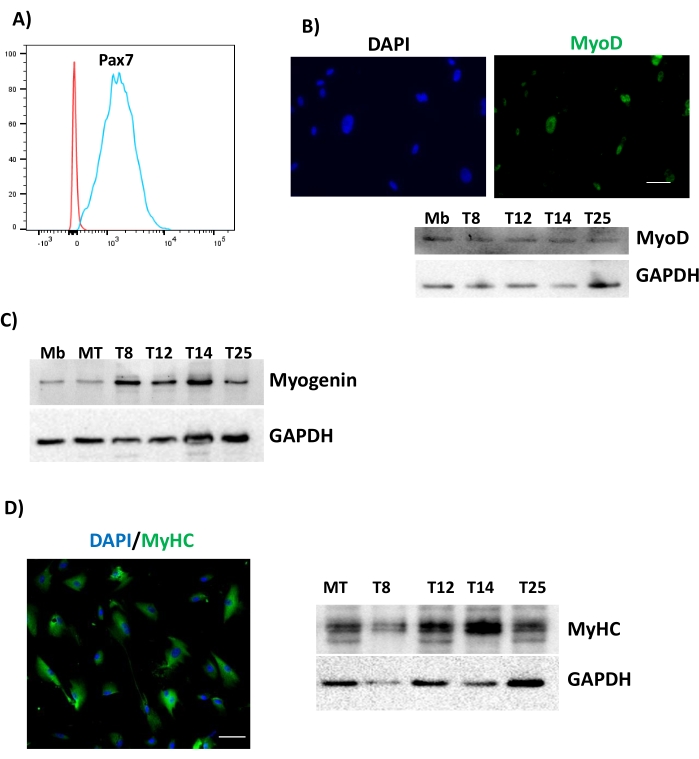
Figure 3: Differentiation of uMSCs into skeletal muscle. uMSCs were cultured in M1 medium for 2 days, 4 days, 7 days, and 10 days and assessed for (A) Pax7 after 2 days, (B) MyoD after 4 days, (C) Myogenin expression from different uMSC lines, denoted by T8, T12, T14, and T25 after 7 days, and (D) MyHC after 10 days. Scale bars = (B, D) 50 µm. Abbreviations: uMSCs = mesenchymal stem cells from human umbilical cord tissue; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; MyoD = myoblast determination protein 1; MyHC = myosin heavy chain; DAPI = 4',6-diamidino-2-phenylindole; Mb = myoblast; MT = myotube. Please click here to view a larger version of this figure.
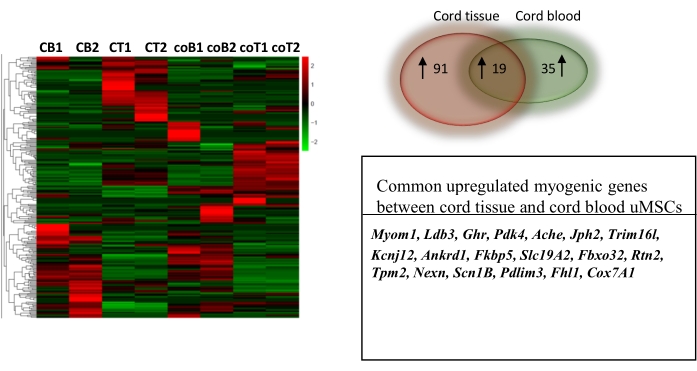
Figure 4: Comparative transcriptomic profiling of uMSCs derived from cord blood and cord tissue differentiated into skeletal muscle. Heat map of normalized counts of 907 genes between control uMSCs and uMSCs differentiated into skeletal muscle for 7 days from cord blood and cord tissue. Venn diagram (left) shows a greater number of myogenic genes upregulated in cord tissue-derived uMSCs compared to cord blood-derived uMSCs. Table below shows the common myogenic genes upregulated in both cord tissue and cord blood. This figure is from Mishra et al.6. Abbreviations: 1, 2 = biological replicates; CB1,2 = myogenic cells derived from uMSCs of cord blood; CT1, 2 = myogenic cells derived from uMSCs of cord tissue; coB1,2 = control undifferentiated uMSCs from cord blood; coT1, 2 = control undifferentiated uMSCs from cord tissue. Please click here to view a larger version of this figure.
For the purposes of conducting a comparative analysis, we compared uMSCs isolated from cord blood and cord tissue. RNA sequencing data revealed that there were more myogenic genes upregulated in uMSCs from cord tissue-derived myogenic cells than from those derived from cord blood (Figure 4). The RNA sequencing data used to support this study is uploaded to NCBI (SRA accession is GSE147114). Briefly, tissue-specific transcriptomic analysis using the PANTHER GO-slim database identified cytoskeletal proteins associated with actin binding and sarcomere assembly (TPM2, LDB3, PDLIM3, FHL1, NEXN, MYOM1) transporters associated with contractile function (RTN2, SLC19A2, ACHE, SCN1B, SLC19A2, JPH2, KCNJ12, ANKRD1), muscle mass maintenance (FBXO32, TRIM16L, GHR), calcium signaling (FKBP5), and enzymatic function (COX7A1, PDK4) (Figure 4). Altogether, these data demonstrate that cord tissue-derived uMSCs represent a compartment displaying robust myogenic potential.
Due to individual variation between uMSC lines that might arise from efficiencies in isolation, heterogeneity within the uMSC compartment, the age of the mother, and the mother's health status, including her nutrient levels, there might be differences in proliferation rates and myogenic potentials between established uMSC lines. However, despite differences in the kinetics of expression, the overall trend of increasing myogenicity with the stagewise expression of myogenic markers is maintained.
Supplemental Figure S1: MSC marker analysis in cord blood-derived uMSCs. uMSCs display the expression of CD105 and hematopoietic markers, CD34 and CD45. Analysis of the CD105+ population shows that only a small proportion of these cells co-express CD90 (N = 5). Abbreviation: uMSCs = mesenchymal stem cells from human umbilical cord tissue. Please click here to download this File.
