Defining adult and larvae population gates
Here, synchronized C. elegans L1s were grown on an NGM plate seeded with E. coli OP50 (Eco), a standard laboratory diet. C. elegans populations were collected for LPS analysis after 96 h or 120 h of growth at 20 °C (Figure 2A). A dot plot of extinction (EXT, a proxy of body density) versus time-of-flight (TOF, a proxy of body length) creates two visually separated clouds of animals. Each dot represents a single animal where higher EXT and TOF values are observed from adults compared to larvae (Figure 2B). These two parameters are valuable inferences for population growth and physiology. For example, a density plot of larvae TOF can visualize the distribution of larval stages. Progenies from 2-day-old adults were dominated by L1 and L2 stages with a TOF below 200, while most progenies from 3-day-old adults reached L3 and L4 stages (Figure 2C). Additionally, 2D density and box-whisker plots are useful to visualize changes in adult body size and density since values of TOF and EXT increase on Day 3 compared to Day 2 when grown on E. coli (Figure 2D–F). This relationship is typically a linear one during adulthood, but some changes in physiology may impact one feature more than another (e.g., adults without eggs may have lower EXT values without affecting TOF).
Profiling gut microbiome composition using fluorescently tagged microbes
In order to illustrate different levels of colonization, we compare a dominant colonizer of the natural C. elegans microbiome dTomato-tagged Ochrobactrum BH3 (Och) and green fluorescent protein (GFP)-tagged E. coli OP50. These two were seeded individually and in an equal mixture based on OD (1:1 mix) on the NGM plate. Synchronized C. elegans L1s were grown on the three conditions and collected at 120 h to examine colonization dynamics in 3-day-old adults (Figure 3A). 2D density and box-whisker plots showed that there are differences in adult TOF and EXT values when C. elegans is grown on Ochrobactrum BH3 and mix cultures (Figure 3B–E). Gut colonization of bacteria can be inferred by the fluorescence level detected in the individual nematodes. Box-whisker plots of red fluorescent readings show increased Ochrobactrum BH3 colonization in the mix condition than in Ochrobactrum BH3 alone. In contrast, green-fluorescent values indicate lower OP50 colonization in the mix condition than in OP50 alone. Similar trends are observed in TOF-normalized fluorescent signals, which removes the effect of body size and reduces variation within the population (Figure 3F–I). For a defined microbiome with multiple fluorescent-tagged microbes, a dot plot can illustrate colonization patterns for these microbes. For example, in the two-member mix microbiome, a dot plot of red fluorescent protein (RFP) versus GFP channels shows that worms are heavily skewed toward the y-axis (RFP), suggesting OP50 colonization is low in most worms while levels of Ochrobactrum BH3 colonization are evenly distributed in the population (Figure 3J). Similarly, a dot plot of RFP versus EXT can reveal the relationship between Ochrobactrum BH3 colonization levels and host development such as body density (Figure 3K). In addition, the differences in reproduction patterns can be observed by plotting the density plot of the respective larvae population on the three conditions (Figure 3L).
Enrichment for targeted populations based on microbiome colonization
The 3-day-old adults grown on the two-member mix microbiome exhibit a wide range of RFP intensity, indicating individual variations in Ochrobactrum BH3 colonization within the group (Figure 4A). To further separate these sub-groups, sorting gates for high and low RFP were manually drawn to sort 15 individuals from each gate into a 96-well plate, as shown by the RFP image of the whole well (Figure 4B). Under higher magnification, overlay images of bright fields and RFP channels confirm that the sorting method selects high and low Ochrobactrum BH3-colonized worms, which allows for further characterization of phenotypic consequences and molecular drivers (Figure 4C).
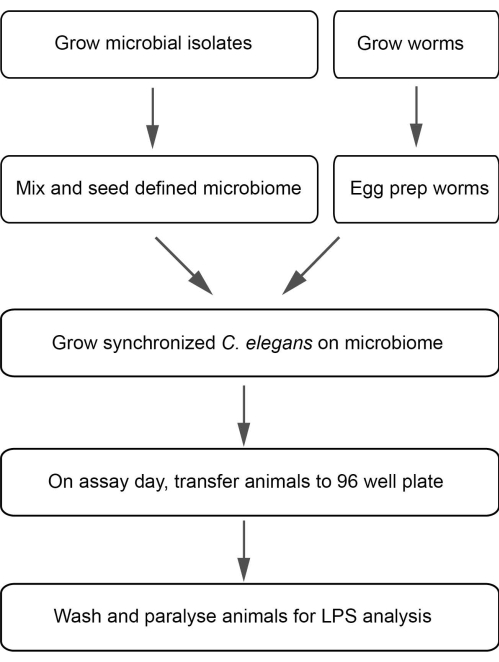
Figure 1: A flow chart for flow vermimetry-based methods to assess gut microbiome and host physiology. Please click here to view a larger version of this figure.
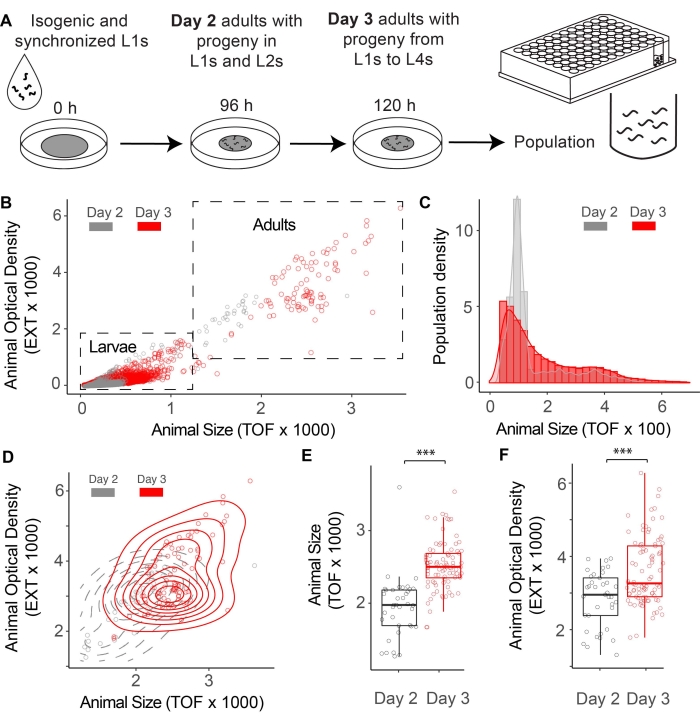
Figure 2: Defining adult and larvae populations. (A) A workflow to collect C. elegans populations on Day 2 and Day 3 of adulthood. (B) Dot plot of time-of-flight (TOF) versus extinction (EXT) for C. elegans populations on Day 2 (Gray) and Day 3 (Red) of adulthood. (C) Density plot of larvae TOF on Day 2 and Day 3 of adulthood. (D–E) Dot and box-and-whisker plots of TOF versus EXT for adults on Day 2 and Day 3 of adulthood. (F) Box-and-whisker plot of adult extinction on Day 2 and Day 3 of adulthood. P-values were calculated with the student's t-test (*** p < 0.001; Day 2 n = 38; Day 3 n = 88). Please click here to view a larger version of this figure.
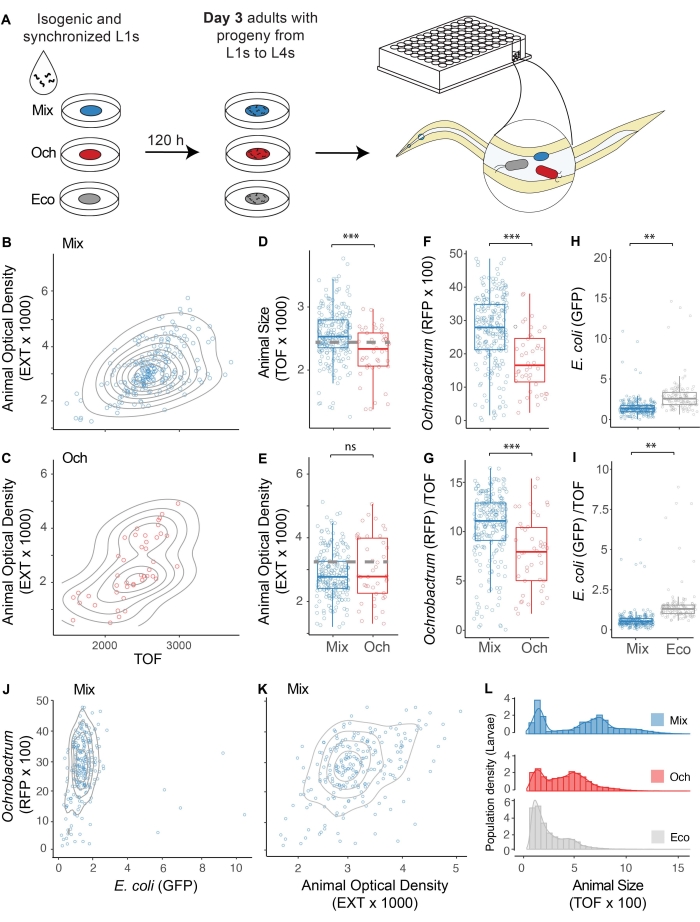
Figure 3: Profiling gut microbiome colonization using fluorescent-tagged microbes. (A) Collection of 3-day-old adult populations grown on Ochrobactrum BH3 (dTomato-expressing; Och), E. coli OP50 (GFP-expressing; Eco) or a 1:1 mix of the two bacteria (mix). (B) Dot plot of TOF versus EXT for Day 3 adults grown on the Mix. (C–E) Dot and box-and-whisker plots of TOF versus EXT for 3-day-old adults grown on Ochrobactrum BH3. Gray dash lines indicate the mean value of the population grown on OP50. (F–G) Box-and-whisker plot of raw and TOF-normalized dTomato (RFP) values for 3-day-old adults grown on Mix and Ochrobactrum BH3 alone. (H–I) Box-and-whisker plots of raw and TOF-normalized GFP for 3-day-old adults grown on mix and OP50. (J) Dot plot of Ochrobactrum BH3 (RFP) versus E. coli OP50 (GFP) for 3-day-old adults grown on mix. (K) Dot plot of RFP versus EXT for 3-day-old adults grown on mix. (L) Density plot of larvae from 3-day-old adults grown on mix, Ochrobactrum BH3, and OP50. All P-values were generated from student's t-test (*** p < 0.001; ** p < 0.01; n.s., not significant; mix n = 230; Och n = 45). Please click here to view a larger version of this figure.
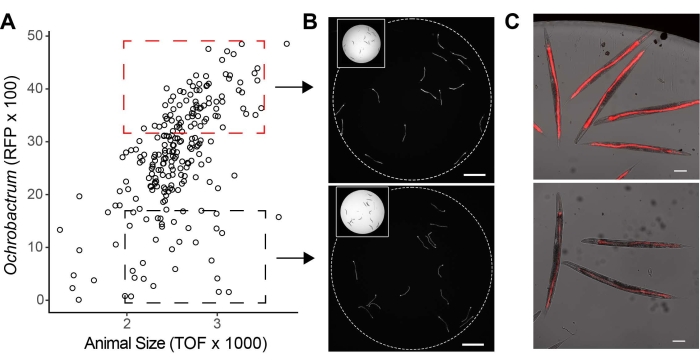
Figure 4: Enrich targeted populations based on microbiome colonization. (A) Dot plot of TOF versus RFP (Ochrobactrum BH3) for 3-day-old adults. High (red box) and low (black box) RFP gates are drawn to enrich C. elegans with high and low Ochrobactrum BH3 colonization. (B) Representative RFP images (4x) of the wells containing 15 sorted worms from high and low RFP gates (Bar = 1 mm). (C) Representative images (10x) of individual worms from high and low RFP gates (Bar = 100 µm). Please click here to view a larger version of this figure.
Supplementary File 1: Representative dataset generated by large particle sorter for N2 populations in Day 2 and Day 3 adulthoods grown on E. coli OP50 and Ochrobactrum BH3. Please click here to download this File.
Supplementary File 2: Scripts used in the analysis and figure generation of representative dataset in the R environment. Please click here to download this File.
