A simple protocol is reported here for obtaining transgenic P. lanceolata plants using A. tumefaciens-mediated transformation. The reporter gene GUS (encoding β-glucuronidase) is transformed, driven by the phloem-expressed promoter of AtPP2, into 3-week-old P. lanceolata roots through A. tumefaciens strain GV3101 (Figure 2). A phloem-specific promoter was chosen because our main interest was to establish a system for the functional genomics of plant vascular tissues, particularly phloem. The method was tested on the root, leaf, and petiole tissue in the preliminary experiment. Although callus could be induced in all tissue types, only the root tissue produced shoot initials (Figure 5A) after 1 month in SIM; the leaf and petiole turned brown and died (Figure 5B). This led to the conclusion that root tissue was the optimal tissue type for use in the transformation method. The roots were incubated in the prepared bacteria resuspended in suspension solution (SS) (Table 1) for a minimum of 20 min, then incubated at room temperature on solid SS plates for up to 3 days in the dark (Figure 3E). The roots were then transferred to the shoot induction medium (SIM) and kept under a grow light, in the conditions indicated in the protocol (step 1.6). Figure 1 and Figure 3 show representative images of each step of the protocol for reference.
Figure 6 shows the progression of shoot initials emerging from transformed tissue, from the first day the roots were placed on the SIM (Figure 6A) to when the shoots were ready to be rooted (Figure 6D). After 1 week, the root tissue formed callus (Figure 6B), and the beginnings of shoot initials could be observed (Figure 6B1). Shoots continued to emerge during weeks 2 and 3 (Figure 6C), and after 4 weeks, the shoots were ready to be transferred to the root induction medium (Figure 6D).
Identification of the putative transgenic plants was conducted using the β-glucuronidase (GUS) histochemical assay, using leaf segments taken once the shoots were around 0.5 cm long. Positive transgenic plants showed the expected staining pattern in the phloem localized tissue, demonstrated in Figure 4. Positive GUS-stained shoots were transferred to the root induction medium, in which they developed robust rooting systems after 4 weeks (Figure 1E). Rooted plants were then transferred to the soil. Figure 4 shows the result of staining in a narrowleaf plantain transformed with the AtPP2 promoter and the β-glucuronidase (GUS) gene, along with a wild-type and a narrowleaf plantain transformed with the AtPP2 promoter, for comparison. All shoots that emerged were confirmed as transgenic. The transformation efficiency was determined to be an average of 20%, with approximately two shoots emerging for every 10 roots that were transformed. Confirmed transgenic plants were transferred to larger pots and grown for 4-8 weeks until they reached the adult stage (Figure 1F).
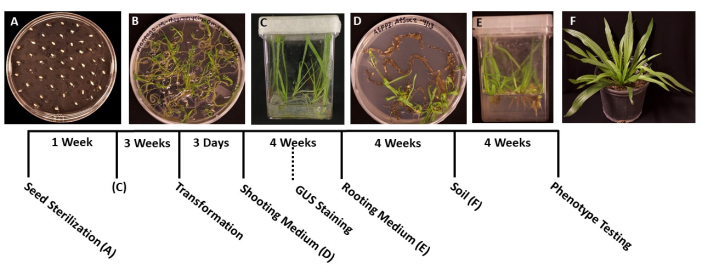
Figure 1: Timeline of Plantago lanceolata transformation. Representative images of each stage of the protocol. (A) Ungerminated seeds plated on an MS plate. (B) Seeds germinated after 1 week, ready to be transferred into magenta boxes. (C) Plants in MS boxes after 3 weeks of growth. Roots are green and healthy, at the ideal stage for transformation. (D) Shoots in shoot induction media after 4 weeks are ready to be transferred into the rooting medium. At this stage, β-glucuronidase (GUS) histochemical staining can be conducted, if applicable. (E) Plants in boxes with root induction media, where roots have formed after 4 weeks of growth. (F) Transgenic plants are grown to full length after 4 weeks of growth in soil. Please click here to view a larger version of this figure.
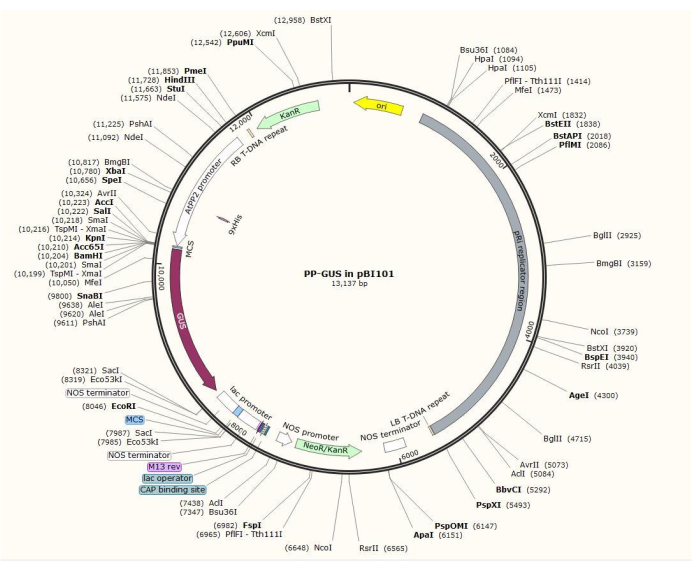
Figure 2: Diagram of the binary vector plasmid pBI101 + β-glucuronidase (GUS) with the inserted phloem-specific promoter AtPP2. Please click here to view a larger version of this figure.
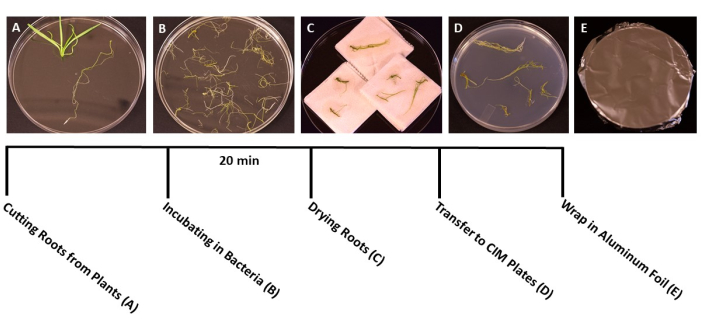
Figure 3: Steps of transformation. Representative images of each step of transformation. (A) Separating roots from shoots during transformation. (B) Soaking roots in bacteria/SS suspension. (C) Drying roots on paper towels to remove excess bacteria. (D) Roots plated on co-culture medium. (E) SS plates wrapped in aluminum foil. Plants were incubated for 2-3 days before being transferred to the shooting medium. Please click here to view a larger version of this figure.
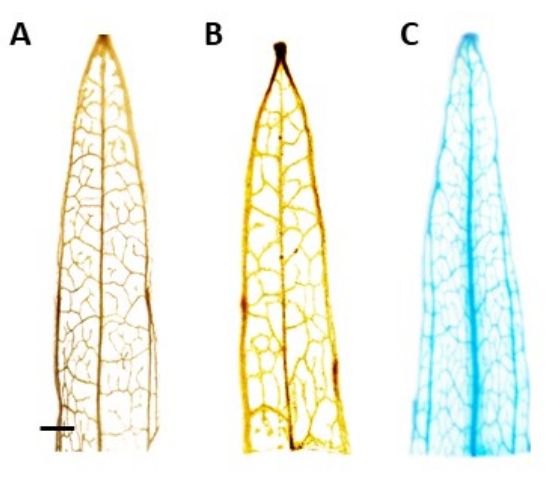
Figure 4: GUS-staining. β-glucuronidase (GUS) staining results of narrowleaf plantain leaf segments. (A) Wild type. (B) Narrowleaf plantain transformed with the plasmid that harbors the AtPP2 promoter (empty vector). (C) Narrowleaf plantain transformed with the plasmid that harbors the AtPP2 promoter and the β-glucuronidase (GUS) gene. Each leaf was stained using the GUS histochemical staining protocol, then imaged with a microscopic camera. Images (B) and (C) show no staining pattern due to the absence of the GUS gene. The right image shows a clear blue staining pattern in the veins, confirming that the plants are transgenic. The bar represents 1 mm, with each leaf segment measuring approximately 1 cm in length. Please click here to view a larger version of this figure.
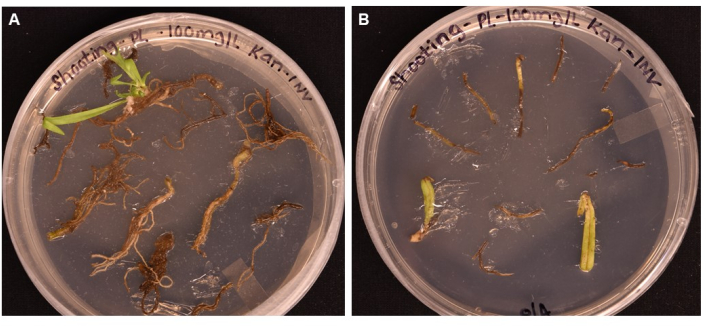
Figure 5: Comparison of transformation efficiency of different tissue types after >1 month incubation on shooting media. (A) Root tissues after over 1 month of growth. Roots have experienced expanded callus, and shoot initials have emerged. Non-transformed callus has begun to die in response to antibiotic selection. (B) Leaf and petiole tissues after over 1 month of growth. Tissues experienced some callus expansion but soon died in response to the antibiotic. No shoots emerged from either tissue. Please click here to view a larger version of this figure.
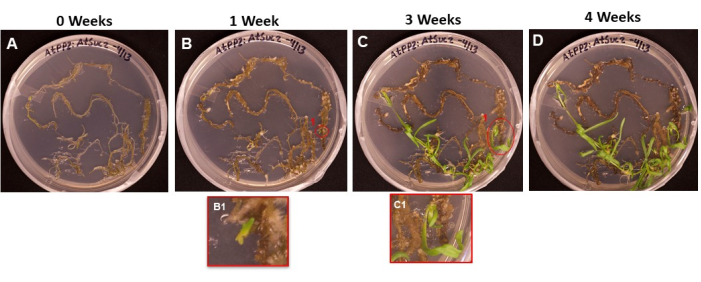
Figure 6: Emergence of callus and shoots on transformed tissue. Representative images of tissues placed on shooting medium after different lengths of incubation. (A) Root tissues just after being plated on the shooting medium. (B) Root tissues after 1 week on shooting medium. Callus expansion can be observed, and (B1) the first shoot initials have begun to emerge. (C) Root tissues after 3 weeks on shooting medium. More shoot initials have emerged. (C1) The shoot that emerged from the B1 shoot initial. (D) Root tissues after 4 weeks of incubation. Non-transformed tissue has begun to turn black/brown and die, and emerging shoots continue to grow. At this stage, shoots are ready to be moved to the rooting medium. Please click here to view a larger version of this figure.
Table 1: Media preparation recipes. A description of how to prepare mediums for transformation. The quantity of vitamins added is calculated based on the indicated stock solution concentration. See Table 2 for vitamin stock solution preparation. For all mediums, add reagents to 900 mL of double-distilled H2O, pH to the indicated level, and then add water to a final volume of 1,000 mL. * = add after sterilization. ** = pH with 1 M KOH. *** = pH with 1 M NaOH. Please click here to download this Table.
Table 2: Vitamin stocks for Plantago mediums. All vitamins must be filter sterilized and labeled accurately before storage. Where indicated, dissolve the powders first in 1 N NaOH, then make up the desired volume with double distilled H2O. Please click here to download this Table.
