MNNG induces PLGC progression in animal model and promotes morphological transformation of GES-1 cells
Upon macroscopic observation, the gastric mucosa of rats in the control group appeared uniformly bright red, smooth, and soft, with the mucosal folds arranged in a linear pattern. In contrast to the rats in control group, the gastric mucosa of the rats in model group exhibited paleness and roughness, and the mucosal plicas were flat or even disappeared (Figure 1B). Starting from the 12th week, histological examination was employed to observe the pathological changes in the gastric mucosa of rats. In the 12th week, histological examination revealed inflammation and vasodilation in the gastric mucosa, along with slight defects in the epithelial cells. The mucosal thickness was normal, and the glands were regularly arranged. In the 16th week, the gastric mucosa became significantly thinner, and the gastric glands were reduced and vacuolated. IM and Dys were occasionally observed with extensive cystic dilatation of the gastric glands at week 20. Until the 24th week, Dys was observed in the gastric mucosa. The intrinsic glands exhibited a decrease in number and were abnormally aligned. Cytological atypia was also observed, including hyperchromatic nuclei, abnormally large nuclear/cytoplasmic ratio, and loss of polarity (Figure 1A). In vitro, the cell model was successfully constructed by administering MNNG. Some morphological changes were observed in the MC cell model. GES-1 cells were observed to exhibit monolayer growth, appearing as uniform spindle-shaped cells with oval or round nuclei. In contrast, MC cells displayed multi-layer growth and uneven distribution. MC cells were elongated with irregular nuclei and spindle-shape similar to fibroblasts (Figure 1C).
HZJD decoction alleviates the development process of PLGC in vivo and inhibits MC cell proliferation
After verifying the PLGC model, rats were treated for another 10 weeks to further clarify the effect of HZJD decoction. The results of H&E staining indicated that the gastric mucosa in the control group was thick with abundant glands and massive parietal cells. On the contrary, the rats in model group displayed pathological changes, including IM and Dys. The pathology of gastric mucosa was significantly improved in HZJD group compared with the model group. In detail, the gastric mucosa demonstrated inflammation, but the epithelial cells remained relatively intact. The glands displayed a regular arrangement, and the parietal cells were scattered. However, the therapeutic effect of positive drug is not obvious (Figure 2A). To further clarify the degree of PLGC, HID-AB staining and AB-PAS staining were performed for IM classification. HID-AB staining was used to differentiate sulfomucins from sialomucins and AB-PAS staining was used to distinguish acidic mucins from neutral mucins13,22. In the control group, gastric mucosal epithelial cells displayed a uniform distribution of blue color instead of brown in HID-AB staining. Additionally, in AB-PAS staining, the epithelial cells displayed a purplish-red color. The gastric mucosa of model group showed brown color in HID-AB staining and blue in AB-PAS staining, which indicated the presence of colonic-type metaplasia (C-IM). In HZJD group, the gastric mucosa did not exhibit a brown stain in HID-AB staining, but predominantly displayed a purplish-red color in AB-PAS staining (Figure 2B). The staining results revealed that HZJD could inhibit the progression of atypical hyperplasia.
To detect whether HZJD can affect cell viability, CCK-8 assay was used to assess the effect of HZJD on MC cells. The cells were treated at different time courses of 24 h, 48 h, and 72 h. The results of CCK-8 assay showed that there was no significant difference in cell viability among groups at 24 h (p > 0.05). At 48 h, the viability of MC cells was significantly increased, compared with GES-1 (p < 0.0001). HZJD decoction inhibited cell viability (p < 0.0001). Although the cells in positive drug group displayed lower cell viability compared with MC, this decrease was not statistically significant (p = 0.1681). The results of CCK-8 assay indicated that HZJD treatment inhibited MC viability in a time-dependent manner (Figure 2D). The evaluation of cell proliferation using the EdU assay yielded results consistent with the findings of the CCK-8 assay. Notably, the cell proliferation of MC cells was significantly higher compared to that of GES-1 cells (p < 0.0001). In HZJD group, the ratio of EdU was obviously decreased, compared with MC cells (p < 0.001). The above results indicated that HZJD had an inhibitory effect on MC cell proliferation (Figure 2C,E).
HZJD decoction promotes mitophagy in PLGC rats and MC cells
To explore the involvement of mitophagy in PLGC, we detected the expression levels of mitophagy-related molecules, including Parkin, Tomm20, P62 and LC3 II/I. In vivo, the protein expression levels of parkin and LC3 II/I were obviously decreased in model rats, compared with control group. Treatment with HZJD decoction led to a marked increase in the protein levels of Parkin and LC3 II/I. The protein expression levels of Tomm20 and P62 in model rats were conspicuously elevated compared with those in control group, whereas HZJD treatment significantly inhibited the levels of Tomm20 and P62 (Figure 3A,C). At the same time, the protein expression levels of Parkin, Tomm20, P62 and LC3 II/I were detected in vitro. The results showed that the protein expression levels of Parkin and LC3 II/I were remarkably down-regulated in MC cells compared to GES-1. HZJD could obviously increase the protein expression levels of Parkin and LC3 II/I. MC cells displayed an increased expression of Tomm20 and P62 proteins, whereas HZJD treatment reduced the excessive expression (Figure 4A,C).
The mRNA levels of Parkin, Tomm20, P62, and LC3 II/I were also examined by RT-qPCR both in vivo and in vitro. The results demonstrated that the mRNA expression levels of parkin and LC3 II/I were remarkably diminished in the model rats and MC cells, while those of Tomm20 and P62 were significantly elevated compared with GES-1 cells and control rats. The gastric tissues and cells treated with HZJD showed upregulation of Parkin and LC3 II/I, while down-regulation of Tomm20 and P62 (Figure 3B, Figure 4B). Another noteworthy result in the animal experiment was that treatment with the positive drug reversed the changes in the mRNA levels of Parkin, LC3 II/I, and Tomm20 in the model rats, but this regulatory effect did not reach statistical significance (p > 0.05; Figure 3B). These results suggest that the positive drug is considerably less effective than HZJD.
In addition, the immunofluorescence analysis revealed that the signals of COX IV and LC3 in the model rats were reduced compared with the control rats. HZJD markedly increased the expression levels of COX IV and LC3 (Figure 3D,E), suggesting that HZJD could promote mitophagy in the gastric tissues of PLGC rats. To further validate the effects of HZJD on mitophagy in MC cells, we examined the fluorescent intensity of mitophagy by staining both mitochondrial phagosomes and lysosomes. When mitochondrial phagosomes and lysosomes fused, the fluorescent intensity (red) changed from weak fluorescence to strong fluorescence. The findings indicated that there was a weak intensity observed in MC cells, but the signal was significantly enhanced in HZJD treated-cells (Figure 4D,E), indicating that HZJD could activate mitophagy inhibited by MNNG in MC cells.
HZJD decoction regulates Sirt3/Foxo3a/Parkin pathway in PLGC rats and MC cells
It is well-known that the Sirt3/Foxo3a/Parkin pathway plays an important role in activating mitophagic pathway23. Thus, we explored whether HZJD could regulate Sirt3/Foxo3a/Parkin pathway in PLGC. Sirt3, Foxo3a, and Parkin were mainly localized in the cytoplasm. The results of immunohistochemistry displayed that the expression levels of Sirt3, Foxo3a, and Parkin were remarkably diminished in the model rats compared to those in the control rats. HZJD demonstrated a significantly superior effect compared to the positive drug in elevating the expression levels of Sirt3, Foxo3a, and Parkin (Figure 5A,B). Next, the protein and mRNA expression levels of Sirt3 and Foxo3a were determined both in vivo and in vitro. The results of Western blotting showed that Sirt3 and Foxo3a were sparsely expressed in the PLGC rats and MC cells, whereas the up-regulated expression levels of the above molecules were observed in the control rats and GES-1 cells. We also found that HZJD could increase the expression levels of Sirt3 and Foxo3a when compared with the model group (Figure 5C,E,F). RT-qPCR results revealed that the mRNA levels of Sirt3 and Foxo3a were reduced in PLGC rats and MC cells compared to control rats and GES-1 cells. After HZJD administration, we noted an obvious increasing trend (Figure 5D,G). These results suggest that HZJD can promote mitophagy both in vivo and in vitro, possibly by interfering with Sirt3/Foxo3a/Parkin pathway to some degree.
HZJD promotes mitophagy by activating Sirt3/Foxo3a/Parkin pathway in MC cells
To further elucidate the possible mechanism of HZJD in triggering mitophagy, MC cells were transfected with si-sirt3. As shown in Figure 6, the protein expression levels of Sirt3 and Foxo3a in si-Sirt3 group were significantly decreased compared with those in si-NC group. Then, the protein expression levels of mitophagy-related molecules Parkin, Tomm20, P62, and LC3 II/I were detected. In contrast to MC group, the expression levels of Parkin and LC3 II/I were markedly decreased in si-Sirt3 group, while those of Tomm20 and P62 proteins were over-expressed in si-Sirt3 group. This data shows that the knockdown of sirt3 can inhibit mitophagy in MC cells. We also observed a decrease in the expression levels of Sirt3 and Foxo3a in si-Sirt3+HZJD group compared to HZJD group. Meanwhile, compared with HZJD group, the protein expression of Parkin and the ratio of LC3 II/I were obviously down-regulated, whereas the expression levels of Tomm20 and P62 were also significantly up-regulated in si-Sirt3+HZJD group (Figure 6A,B). This data indicates that si-sirt3 can attenuate mitophagy caused by HZJD decoction. The results of RT-qPCR were in good agreement with those Western blotting (Figure 6C). In addition, we also observed the ultrastructure of MC cells using transmission electron microscopy. The results showed that the mitochondrial structures of GES-1 cells were basically normal without obvious mitochondrial damage. Moreover, the swollen mitochondrion in MC cells was observed and the mitochondrial cristae disappeared, indicating that mitophagy in MC cells was reduced. In si-Sirt3 group, swollen mitochondrial membranes were observed, and the degree of mitochondrial disruption was increased compared with si-NC group. We also observed an accumulation of autophagic vesicles in HZJD group when compared to si-Sirt3+HZJD group (Figure 6D). All these results further confirm that HZJD decoction induces mitophagy by regulating Sirt3/Foxo3a/Parkin pathway.
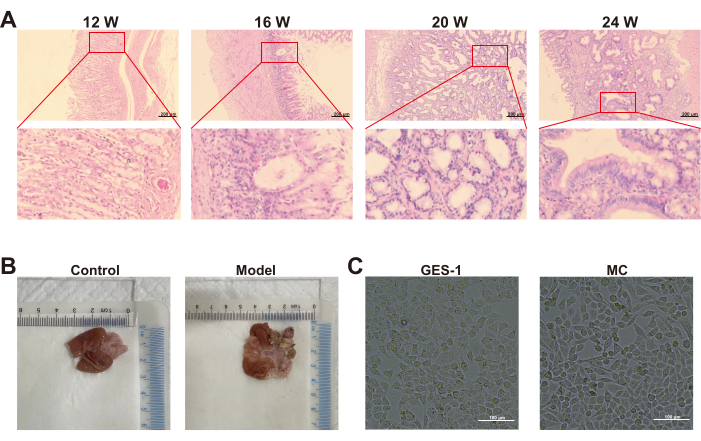
Figure 1: MNNG induces morphologic and pathologic changes in animal model and GES- 1 cells. (A) The 12th, 16th, 20th, and 24th week pathology observations of model group (H&E staining, 100x magnification). (B) The 24th week morphology of gastric mucosa in control group and model group (n=6). (C) The cell morphology in GES-1 group and MC group (n=3). Abbreviations: 12 W = 12 week, 16 W = 16 week, 20 W = 20 week, 24 W = 24 week. Please click here to view a larger version of this figure.
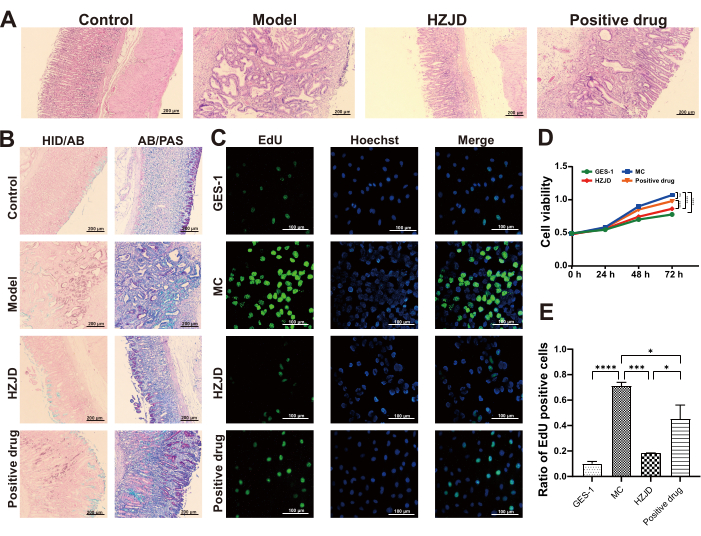
Figure 2: HZJD decoction improves gastric mucosal histopathology and inhibits MC cells proliferation. (A) Representative pathological images of gastric tissues in each group (H&E staining, 100x magnification; n=6). (B) Representative images of gastric tissues in each group (HID/AB staining and AB/PAS staining, 100x magnification, n=6). (C) Cell proliferation levels in each group are tested by EdU assay (n=6). (D) Cell viability levels in each group, as tested by CCK-8 assay (n=6). (E) Quantitative analysis of cell proliferation. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, nsp>0.05. Please click here to view a larger version of this figure.
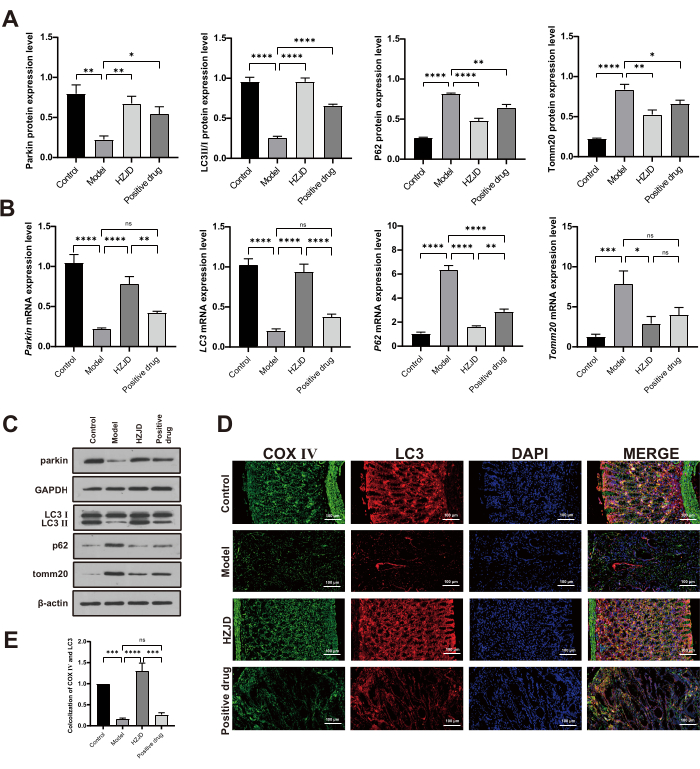
Figure 3: HZJD decoction improves mitophagy in PLGC rats. (A) Data summary for the relative protein expression levels of Parkin after normalization to GAPDH, as well as LC3 II/I, P62 and Tomm20 after normalization to β-actin (n=3). (B) Data summary for the mRNA expression levels of Parkin, LC3, P62 and Tomm20 (n=9). (C) The protein expression levels of Parkin, LC3 II/I, P62 and Tomm20 (n=3). (D, E) The fluorescence co-localization analysis of COX IV and LC3 (n=6). *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, nsp>0.05. Please click here to view a larger version of this figure.
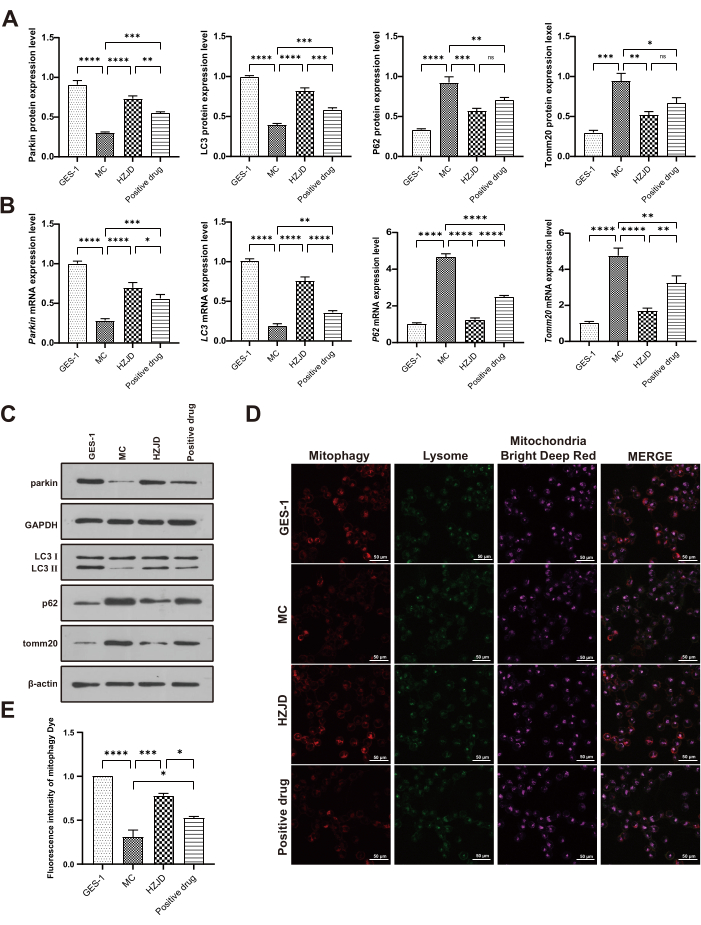
Figure 4: HZJD decoction improves mitophagy in MC cells. (A) Data summary for the relative protein expression levels of Parkin after normalization to GAPDH, as well as LC3 II/I, P62 and Tomm20 after normalization to β-actin (n=3). (B) Data summary for the mRNA expression levels of Parkin, LC3 II/I, P62, Tomm20 (n=9). (C) The protein expression levels of Parkin, LC3 II/I, P62 and Tomm20 (n=3). (D, E) Representative confocal images of cell mitophagy in each group (n=3). *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, nsp>0.05. Please click here to view a larger version of this figure.
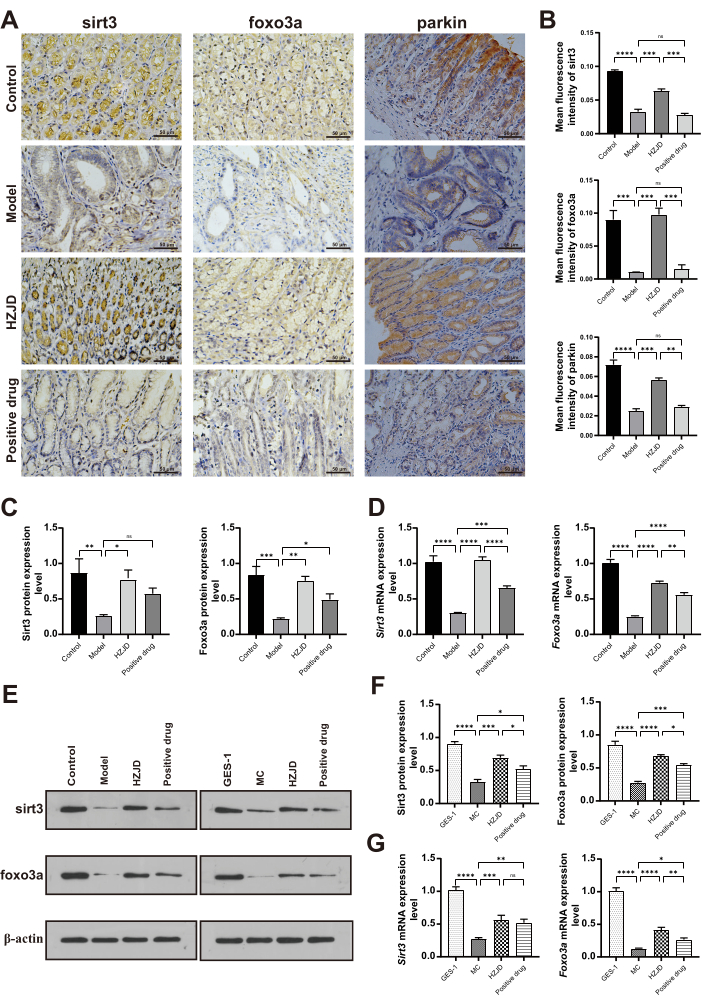
Figure 5: HZJD decoction regulates Sirt3/Foxo3a/Parkin pathway both in vivo and in vitro. (A, B) Representative immunohistochemical images of Sirt3, Foxo3a, and Parkin in gastric tissues in each group (n=6). (C, E) The protein expression levels of Sirt3 and Foxo3a in vivo (n=3). (D) The mRNA expression levels of Sirt3 and Foxo3ain vivo (n=9). (F) The protein expression levels of Sirt3 and Foxo3a in vitro (n=3). (G) The mRNA expression levels of Sirt3 and Foxo3ain vitro (n=9). *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, nsp>0.05. Please click here to view a larger version of this figure.
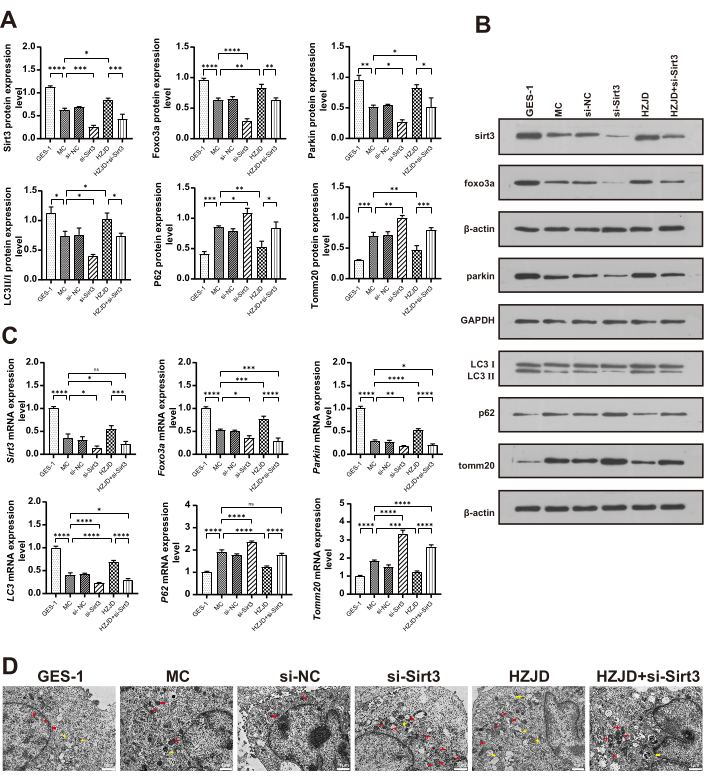
Figure 6: si-sirt3 reverses the effect of HZJD decoction on PLGC cell model. (A, B) The protein expression levels of Sirt3, Foxo3a, Parkin, LC3 II/I, P62, and Tomm20 in each group in vitro (n=3). (C) The mRNA expression levels of Sirt3, Foxo3a, Parkin, LC3, P62, and Tomm20 in each group in vitro (n=9). (D) Representative TEM images of cellular ultrastructure in each group. Red arrows represent mitochondrion. Yellow arrows represent autophagic vacuoles (n=3). *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, nsp>0.05. Here, si-NC represents negative control group, si-Sirt3 represents Sirt3 silencing group, si-Sirt3+HZJD represents Sirt3 silencing combined with HZJD decoction group (si-Sirt3+HZJD). Please click here to view a larger version of this figure.
| Chinese name | Latin name | Ratio |
| Yinchen | Artemisia capillaris Thunb. | 11% |
| Baihuasheshecao | Hedyotis diffusa Willd. | 11% |
| Banlagen | Isatis tinctoria Linnaeus. | 11% |
| Banbianlian | Lobelia chinensis Lour. | 11% |
| Banzhilian | Scutellaria barbata D. Don. | 11% |
| Jiaogulan | Gynostemma pentaphyllum (Thunb.) Makino. | 11% |
| Huangqin | Scutellaria baicalensis Georgi. | 8% |
| Huanglian | Coptis chinensis Franch. | 8% |
| Kushen | Sophora flavescens Alt. | 7% |
| Guanghuoxiang | Pogostemon cablin (Blanco) Benth. | 6% |
| Peilan | Eupatorium fortunei Turcz. | 6% |
Table 1: Drug Composition and proportion contained in HZJD decoction.
| Genes | Forwad primer (5’-3’) | Reverse primer (5’-3’) |
| Sirt3 | GGCGGCAGGGACGATTATTA | AAGGGCTTGGGGTTGTGAAA |
| Foxo3a | AAGGGAGGAGGAGGAATGTGGAA | GGTTGTGCCGGATGGAGTT |
| Parkin | AGGATTACCCAGGAGACCGC | ATCATGGTCACTGGGTAGGTG |
| P62 | TGAGAGACAAAGCCAAGGAGG | CTGGTCCCATTCCAGTCGTC |
| LC3 | CAGCGTCTCCACACCAATCT | GCGGGTTTTGTGAACCTGAA |
| Tomm20 | ATGAGCTGGGTGTCACTTGG | GTGCATTCTTCTGGCAGGGA |
| β-actin | CCGTTGCCCTGAGGCTCTTT | CCTTCTGCATCCTGTCAGCAA |
Table 2: Primer sequences used in RT-qPCR analysis.
| 1-Methyl-3-nitro-1-nitrosoguanidine | Shanghai McLean Biochemical Technology Co., Ltd., Shanghai, CHN | R030453 | |
| 3,3’-diaminobenzidine | Beijing Zhong Shan-Golden Bridge Biological Technology Co., Ltd., Beijing, CHN | ZLI0919 | |
| Alcian blue periodic acid schiff staining reagent kit | Beijing Solarbio Technology Co., Ltd., Beijing, CHN | G1285 | |
| Anti-fade fluorescence mounting medium | Beijing Solarbio Technology Co., Ltd., Beijing, CHN | S2110 | |
| Bicinchoninic acid protein concentration determination kit | Wuhan Servicebio Technology Co., Ltd., Wuhan, CHN | G2026-1000T | |
| CCK-8 reagent kit | Boster Biological Engineering co., Ltd., Wuhan, CHN | AR1160 | |
| Confocal fluorescence microscopy | Leica Instruments Co., Ltd., Weztlar, GER | TCS-SP8SR | |
COX  antibody antibody |
Abcam Trading Co., Ltd., Shanghai, CHN | ab202554 | |
| DAPI staining solution | Beijing Solarbio Technology Co., Ltd., Beijing, CHN | C0065 | |
| DEPC | Wuhan Servicebio Technology Co., Ltd., Wuhan, CHN | G3004 | |
| EdU cell proliferation kit | Beyotime Biotechnology Co., Ltd., Shanghai, CHN | C0071S | |
| Electrophoresis solution | Boster Biological Technology co., Ltd., Wuhan, CHN | AR0139 | |
| Ethanol | Tianjin Baishi Chemical Industry Co., Ltd., Tianjin, CHN | 64-17-5 | |
| FBS | Gibco Corporation, Gaithersburg, USA | 16000044 | |
| Fluorescence microscope | Olympus Corporation, Tokyo, JPN | BH2-RFCA | |
| Foxo3a antibody | Abcam Trading Co., Ltd., Shanghai, CHN | 23683 | |
| GAPDH antibody | Wuhan Sanying Biology Technology Co., Ltd., Wuhan, CHN | 10494-1-AP | |
| Gel preparation kit | Boster Biological Technology co., Ltd., Wuhan, CHN | AR0138 | |
| GES-1 cell | Procell Life Science&Technology Co., Ltd., Wuhan, CHN | CL-0563 | |
| Goat-anti-mouse IgG | CST Biological Reagents Co., Ltd., Shanghai, CHN | 4409 | |
| Goat-anti-rabbit IgG | Abcam Trading Co., Ltd., Shanghai, CHN | ab150077 | |
| Hematoxylin-eosin staining solution | Zhuhai Beso Biotechnology Co., Ltd., Shenzhen, CHN | BA4027 | |
| High iron diamine/alcian blue staining reagent kit | Beijing Solarbio Technology Co., Ltd., Beijing, CHN | G2070 | |
| ImageJ | National Institutes of Health, Bethesda, USA | ||
| Image-Pro Plus 6.0 software | Media Cybernetics Inc., Maryland, USA | ||
| Isopropanol | Tianjin Baishi Chemical Industry Co., Ltd., Tianjin, CHN | 67-63-0 | |
| LC3 antibody | CST Biological Reagents Co., Ltd., Shanghai, CHN | 83506 | |
| Loading buffer | Boster Biological Technology co., Ltd., Wuhan, CHN | AR0198 | |
| Microplate reader | Rayto Life and Analytical Sciences Co., Ltd., Shenzhen, CHN | RT6100 | |
| Microtome | Leica Instruments Co., Ltd., Weztlar, GER | RM2245 | |
| Mitophagy kit | Dojindo Laboratories, Kyushu Island, JPN | MD01-10 | |
| Neutral balsam | Wuhan Servicebio Technology Co., Ltd., Wuhan, CHN | WG10004160 | |
| Optical microscope | Olympus Corporation, Tokyo, JPN | BH2-RFCA | |
| P62 antibody | Abcam Trading Co., Ltd., Shanghai, CHN | ab91526 | |
| Paraformaldehyde | Biosharp Life Sciences, Anhui, CHN | BL539A | |
| Parkin antibody | CST Biological Reagents Co., Ltd., Shanghai, CHN | 32833 | |
| Penicillin–streptomycin | Gibco Corporation, Gaithersburg, USA | 15140122 | |
| phosphate-buffered saline | Wuhan Servicebio Technology Co., Ltd., Wuhan, CHN | G0002-15 | |
| PVDF membrane | Millipore Corporation, Michigan, USA | IPVH00010 | |
| Redzol | SBS Genetech Co., Ltd., Beijing, CHN | FTR-50 | |
| Reverse transcription reagent kit | Igene Biotechnology Co., Ltd., Guangzhou, CHN | QP057 | |
| RIPA Buffer solution | Beijing Solarbio Technology Co., Ltd., Beijing, CHN | R002 | |
| Roswell Park Memorial Institute | Gibco Corporation, Gaithersburg, USA | 11875093 | |
| Silicone needle | Zhongke Life Biotechnology Co., Ltd., Beijing, CHN | TFEP-2 | |
| siRNA | Wuhan Genecreate Biological Engineering Co., Ltd. ,Wuhan,CHN | ||
| Sirt3 antibody | Abcam Trading Co., Ltd., Shanghai, CHN | 189860 | |
| Sodium salicylate | Shanghai Aladdin Biochemical Technology Co., Ltd., Shanghai, CHN | S104176 | |
| Sprague-Dawley rat | Beijing Huafukang Biotechnology Co., Ltd., Beijing, CHN | 110322210102553975 | |
| SYBR quantitative PCR kit | Wuhan Servicebio Technology Co., Ltd., Wuhan, CHN | G3320-15 | |
| Tomm20 antibody | Abcam Trading Co., Ltd., Shanghai, CHN | ab186735 | |
| Transferring buffer | Boster Biological Technology co., Ltd., Wuhan, CHN | AR0141 | |
| Trichloromethane | Shanghai McLean Biochemical Technology Co., Ltd., Shanghai, CHN | 821112 | |
| Tris-buffered saline with Tween 20 | Wuhan Servicebio Technology Co., Ltd., Wuhan, CHN | G0004 | |
| VDAC1 antibody | Abcam Trading Co., Ltd., Shanghai, CHN | ab15895 | |
| Xylene | Tianjin Baishi Chemical Industry Co., Ltd., Tianjin, CHN | 1330-20-7 | |
| β-actin antibody | Abcam Trading Co., Ltd., Shanghai, CHN | ab8226 |
