Kisspeptin-expressing neurons (Kiss1 gene) located in the arcuate nucleus of the hypothalamus are a potent stimulator of GnRH, and thus, of LH release from the pituitary gland24,25. In this protocol demonstration, we have used kisspeptin-induced LH secretion to illustrate the functioning of the automated blood sampling technique. Figure 2 shows representative LH patterns in adult Kiss1-eYFP females that previously received a unilateral stereotaxic injection of AAV-hM3Dq-mCherry in the arcuate nucleus (AP: -4.95, ML: -0.35, DV: -5.7). The ChR2-eYFP was used as a fluorescent reporter for Kiss1 cells. One month after the stereotaxic surgery, the mice underwent carotid artery and jugular vein cannulation and were connected to the automatic blood sampler 4 days post-surgery. Blood collections for determining basal LH levels were started on the next day at 10 min sampling frequency (7 min interval between samples and 3 min/sampling) followed by an automated IV CNO injection and continued blood sampling every 10 min for 30 min. Diestrous LH levels are generally low (Figure 2A), but variations are usually observed because of its pulsatile release (Figure 2B). Following the CNO injection (and activation of kisspeptin neurons), the rise in LH was sharp (within 10 min). The level of increase and the duration of the peak depends on numerous factors, including the site of the injection, the number of neurons activated, or the population targeted.
Figure 3 shows the brain site of injection in the arcuate nucleus of the female mouse represented in Figure 2A. Kiss1-eYFP neurons are labeled in green, while the mCherry immunoreactivity shows the site of the AAV injection and activation following CNO. Activated neurons were detected using cFOS immunoreactivity, labeled with DAB. Most mCherry neurons colocalized with Kiss1-eYFP, and many showed cFOS immunoreactivity, demonstrating that the viral and neuronal activation was specific to the targeted population.
A representative LH pulsatile pattern of release in diestrous wildtype (C57BL/6J) mice followed by the response to an IP injection of kisspeptin-10 is shown in Figure 4. The mouse underwent carotid artery cannulation and was connected to the automatic blood sampling system 4 days after the surgery. The next morning, the estrous cycles were checked and the blood collection and kisspeptin-10 injection were performed on the diestrous day26. Blood samples were collected every 7 min for 2 h (4 min interval, plus 3 min/sampling for 120 min before the injection) to determine the baseline levels and LH pulsatility, followed by an IP injection of kisspeptin-10 (65 µg/kg) and continued blood collection every 7 min for an additional 30 min. Clear LH pulses typical for a female in diestrous were observed, showing low basal LH levels, pulse frequency of ~2 pulses/h, and pulse amplitude ~1 ng/mL27. An immediate and robust increase in LH was detected in response to kisspeptin administration28. The LH secretion patterns and changes after stimulation are in agreement with other studies using manual blood collection10,27,29,30. These results demonstrate that the automated blood sampling method captures typical and stimulated LH secretion under a stress-free condition.
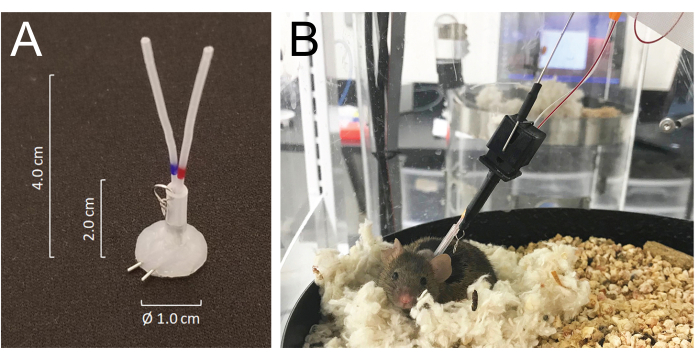
Figure 1: Details of the connector system and mouse connections to the infusion and sampling system tubing. (A) A silicone-coated tubing connector (MASA) constructed using two 25G needle tubing and two PE-20 tubing, sleeved in a silastic tubing attached with a small metal ring. (B) A mouse connected to the infusion and sampling tubing in the sampling cage, resting in its nest during the blood sampling. Please click here to view a larger version of this figure.
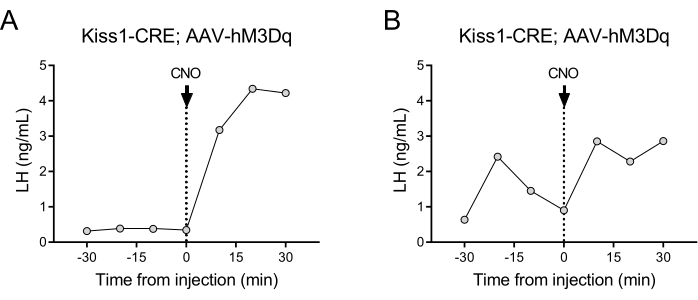
Figure 2: Representative results for LH pulses in Kiss1-Cre female mice injected with AAV-hM3Dq in the arcuate nucleus and remotely activated with CNO. Basal LH levels were measured every 10 min for half an hour. At time 0 following the blood collection, the female received an intravenous injection of clozapine-N-oxide (0.5 mg/kg) and blood continued to be collected from the carotid artery every 10 min for an additional half an hour. (A) Shows a female with low basal LH levels. (B) Shows a female displaying an LH pulse preinjection. Abbreviations: Kiss1 = Kisspeptin; AAV = adeno-associated virus; CNO = clozapine-N-oxide; LH = luteinizing hormone. Please click here to view a larger version of this figure.
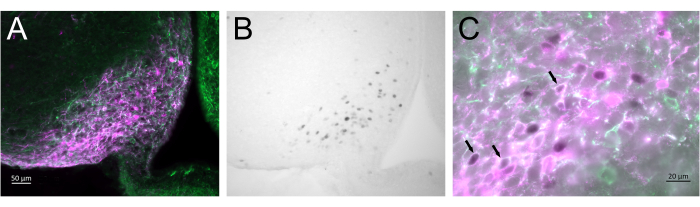
Figure 3: Brain activation in the arcuate nucleus of the Kiss1-Cre;Chr2-eYFP (Kiss1-eYFP) female represented in Figure 2A. ChR2-eYFP was only used as a reporter gene to label kiss1 neurons. (A) Low-magnification fluorescence image showing the site of the AAV injection in the arcuate nucleus. Green: eYFP immunoreactivity, Magenta: mCherry immunoreactivity. (B) Low-magnification brightfield image of the area corresponding to Figure 3A, showing cFOS immunoreactivity (black) in the site of the AAV injection in the Arcuate nucleus. (C) High-magnification fluorescence and brightfield image combined showing a closer look of the neurons in Figure 3A,B. Kiss1-eYFP neurons coexpressing AAV-mCherry that have been activated are those with white cytoplasm and black nucleus (arrows). Scale bars = 50 µm (A,B), 20 µm (C). Abbreviations: Kiss1 = Kisspeptin; AAV = adeno-associated virus; eYFP = enhanced yellow fluorescent protein. Please click here to view a larger version of this figure.
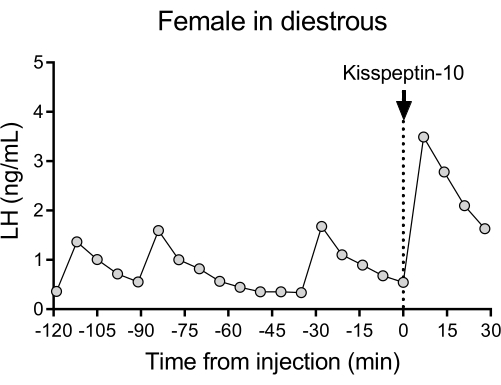
Figure 4: Basal LH pulsatility in a diestrous wildtype female measured every 7 min for 2 h. The female then received an intraperitoneal injection of kisspeptin-10 (65 µg/kg) at time 0, and blood samples were continuously collected every 7 min for half an hour. Abbreviation: LH = luteinizing hormone. Please click here to view a larger version of this figure.
