MAPP was applied to determine the glycan composition of agricultural biomass waste, comprising mango peels from several northern Thai varieties, Coffea arabica cherry pulp and coffee bean processing waste, and root, stem, and leaf tissue from Thai black garlic, Allium sativum var. ophioscorodon. Several plant-derived polysaccharides are used in the food industry as functional ingredients42,43. Thus, the aim of this experiment was to deduce whether these abundant and currently underutilized agro-industrial waste materials may provide a source of value-added pure polysaccharides.
AIR material is routinely employed to prepare samples intended for glycan analysis44. There are several advantages to using AIR; treatment with solvents effectively removes endogenous CAZymes, metabolites, small saccharides, lipids, and pigments, resulting in samples enriched with polysaccharides and structural proteins34. Furthermore, producing AIR is a fast and effective way to increase sample longevity, as it is thermostable and can be stored for several years.
Three mixed fractions of constituent glycans were sequentially extracted from plant AIR material using CDTA, NaOH, and cellulase. CDTA chelates Ca2+ ions, which allow the removal of Ca2+ crosslinked de-esterified pectins from plant cell walls45. Alkaline conditions allow predominantly hemicelluloses, such as mannan, xylan, and β-glucan, to be released due to the disruption of hydrogen bonding and saponification of ester linkages between cellulose microfibrils and hemicellulose, and lignin and hemicellulose, respectively46. A recombinant endo-1,4-β-glucanase from Bacillus spp. was used to degrade amorphous regions of the structural cellulose microfibrils, releasing residual glycans bound to cellulose within the cell walls47. Although this method effectively separates glycans into these three broad groups, it should be noted that the samples are not pure; by the very nature of the extraction method, hemicellulose, if present in the sample, will inevitably be extracted and subsequently detected to varying degrees in the CDTA and cellulase fractions. Likewise, some pectin will be detected in the NaOH extraction if present in the sample.
A non-contact, piezoelectric microarray printing robot was used to immobilize extracted glycan fractions onto nitrocellulose via non-covalent attachment11, forming 300 identical microarrays. Defined glycan standards (Table 1) were also included in the printed microarrays as positive controls (Figure 5). The MAPP binding profile obtained for the selected glycan standards corresponds to previously reported epitope specificities. For example, LM21 exhibited strong binding to multiple mannan polysaccharides (galactomannan and glucomannan), while LM22 exhibited only weak binding to galactomannan25. Similarly, LM19 preferentially bound to de-esterified homogalacturonan48 and LM15 bound to tamarind seed xyloglucan23.
The relative abundance of 16 epitopes, diagnostic of non-cellulosic plant cell wall polysaccharides, were detected by the attachment of glycan-directed monoclonal antibodies (Table 2) to printed extracts (Figure 6). The majority of extracted glycans were detected within the alkaline NaOH fraction. Strong binding signals were recorded for mAbs LM10 and LM11, representing xylan/arabinoxylan, within the peels of all the mango varieties tested. Within the garlic samples, LM10 and LM11 bound preferentially to root tissue extract (Garlic R) and exhibited only weak binding to the leaf tissue extract (Garlic L). LM19, representing partially methyl-esterified or un-esterified homogalacturonan, bound strongly to some mango variety extracts (Aokrong and Talabnak), but bound only weakly, or its binding was undetectable, in other varieties (Chokanan, Mamkamdang, Mahachanok, and Nga). In addition, LM19 bound only to the coffee pulp fractions and did not bind to the coffee bean processing waste material, previously thought to be composed of semi-purified coffee pectin (unpublished data).
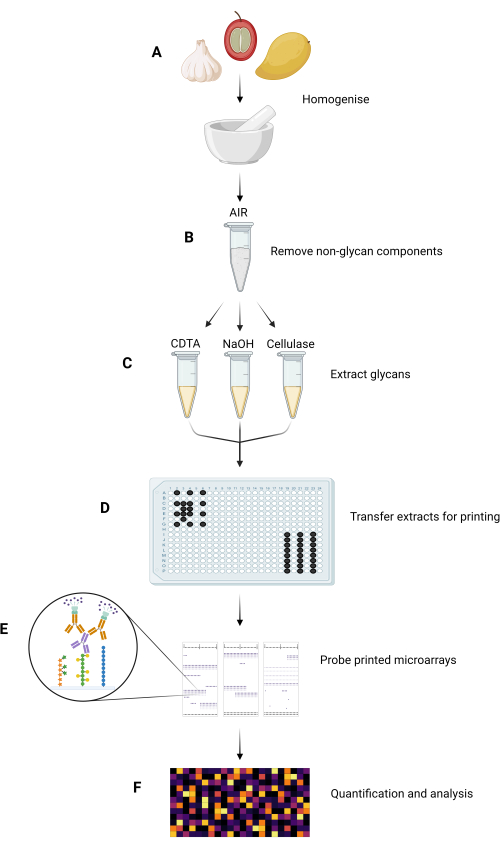
Figure 1: Major experimental steps in the MAPP method. (A) Samples are homogenized to form fine powders. (B) The homogenized samples are processed to isolate their AIRs. (C) The constituent glycans are sequentially extracted using a tailored extraction regime. (D) The extracted glycan fractions, ink, and GSB are transferred into 384-well plates, according to the plate layout, for printing onto nitrocellulose. (E) The printed microarrays are probed with selected GRMPs. (F) GRMP binding to the printed glycan fractions is quantified and analyzed before data are presented as a heatmap. Please click here to view a larger version of this figure.
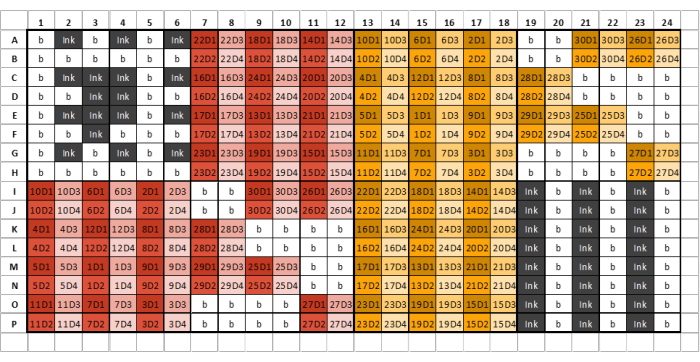
Figure 2: Example of a 384-well plate layout for sample, ink, and GSB loading with four dilutions per extracted glycan sample/standard. Different colors denote samples arising from different extraction reagents, while different shades represent serial dilutions. The first number in the code represents the sample number, while the end number represents the dilution number (D1 denotes dilution one, D2 denotes dilution two, and so on). For example, a well labeled '12D3' represents glycan sample 12, dilution three. Well plates should be divided into eight identical sections comprising six columns and eight rows. The first section of the first plate should contain only ink and buffer and resemble the example plate layout. Extracted glycan samples can then be loaded into subsequent plate sections according to the plate layout. Different extraction reagents should not be loaded into the same plate section. If there are insufficient samples to fill an entire section, fill all remaining wells in that section with buffer; do not leave any wells empty. If multiple plates are required, the next section after all samples have been loaded should contain three alternating columns of ink and GSB-this may not be section eight, depending on the number of samples being printed. Please click here to view a larger version of this figure.
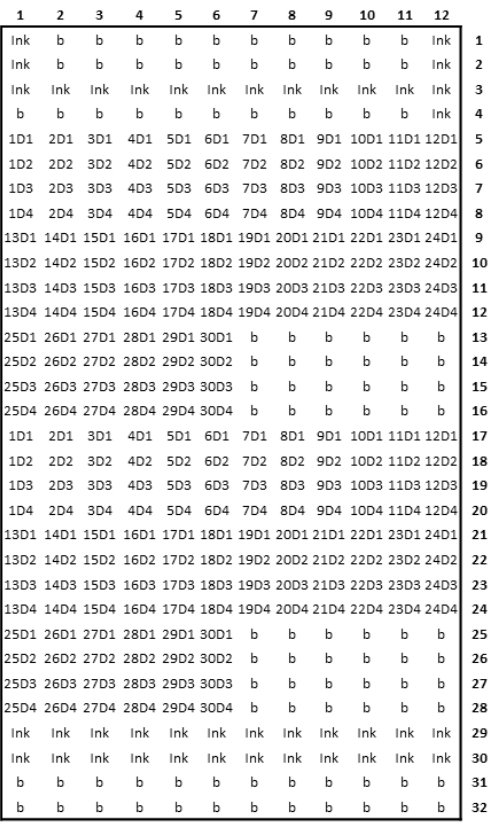
Figure 3: Schematic representation of printed microarray design. Please click here to view a larger version of this figure.
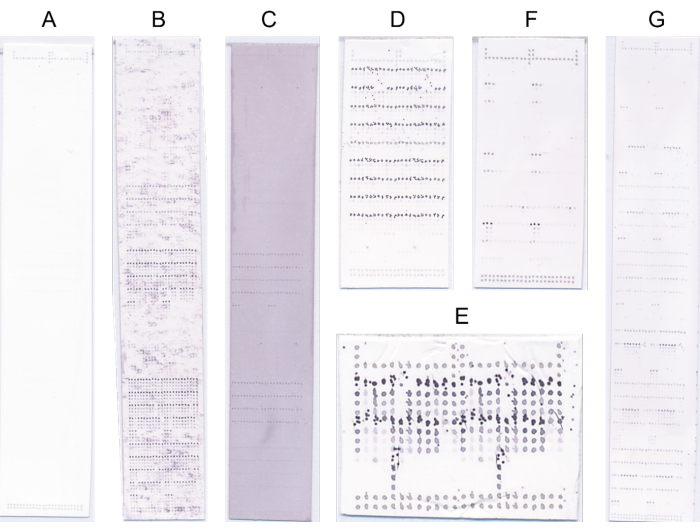
Figure 4: Representative microarrays. (A) No binding. (B) The binding signal is obscured by high background signal. (C) Generalized blue/purple staining due to oversaturation with NBT/BCIP. (D) Defective probing due to high substrate concentration. (E) Defective printing due to the unclean print head. (F) Strong binding to few samples. (G) Strong binding to many samples. Please click here to view a larger version of this figure.
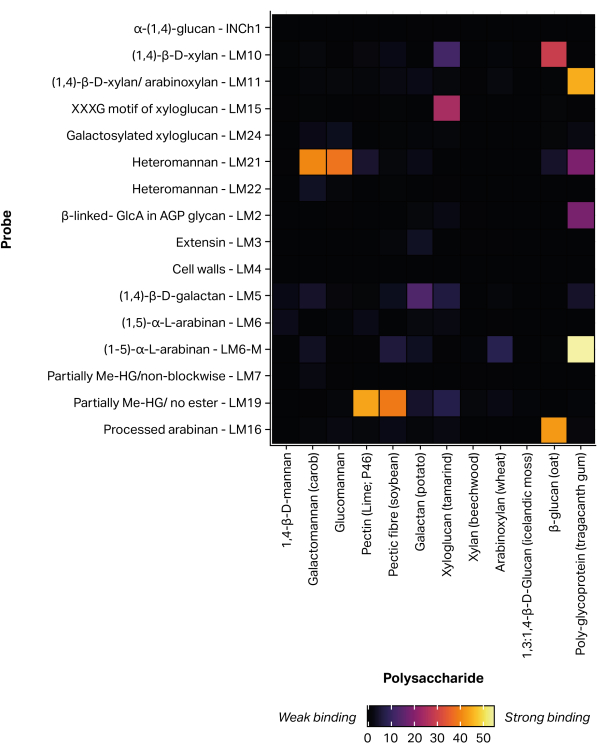
Figure 5: Monoclonal antibody binding to defined glycan standards, included to validate the printing and probing process. Please click here to view a larger version of this figure.
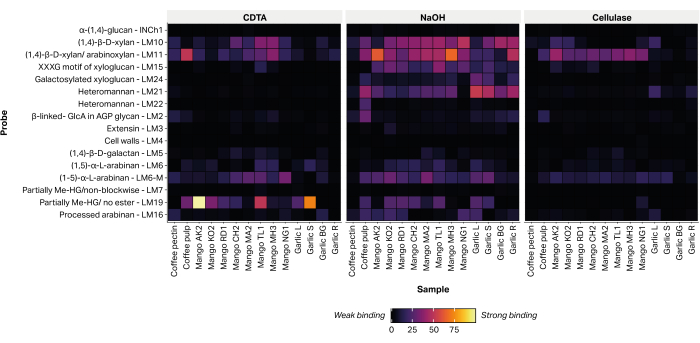
Figure 6: MAPP of glycans extracted from agricultural biomass waste. The samples include coffee pulp waste (coffee pulp and coffee pectin), mango peels from several Thai varieties (AO, Aokrong; KO, Kam; RD, Rad; CH, Chokanan; MA, Mamkamdang; TL, Talabnak; MH, Mahachanok; NG, Nga) and black garlic leaves (Garlic L), stem (Garlic S), bulb (Garlic BG), and roots (Garlic R), using CDTA, NaOH, and cellulase (Bacillus spp. cellulase 5A). Please click here to view a larger version of this figure.
Table 1: Defined commercial polysaccharide standards used in MAPP analysis as positive controls. Please click here to download this Table.
Table 2: Glycan-directed monoclonal antibodies selected for the interrogation of extracted plant glycan microarrays. Please click here to download this Table.
