Pore size measurement, cell distribution, and in vitro mineralization (Figure 1 and Figure 2)
Complete removal of native cellular components of the apple tissue scaffolds was achieved after treating the scaffolds with SDS and CaCl2 (Figure 1A). The scaffolds exhibited a highly porous structure, which was confirmed using confocal microscopy. The quantification of the images demonstrated an average pore size of 154 µm ± 40 µm. The pore size distribution ranged between 73 µm and 288 µm. However, the majority of the pores ranged between 100 µm and 200 µm (Figure 1C).
Following a 4-week culture period in differentiation medium, the cell-seeded scaffolds exhibited widespread white mineral deposits (Figure 1A). The scaffolds containing cells displayed an opaque white coloration, suggesting mineralization, which was not observed in the blank scaffolds (scaffolds without seeded cells). Furthermore, analysis using confocal laser scanning microscopy revealed a homogeneous cell distribution within the scaffolds (Figure 1B).
Scaffolds seeded or not with cells were stained with BCIP/NBT and ARS to analyze ALP activity and mineralization, respectively (Figure 1D). The BCIP/NBT staining revealed a substantial increase in ALP activity (depicted by a strong purple color) within the cell-seeded scaffolds cultured in differentiation medium, in contrast to the blank scaffolds or the cell-seeded scaffolds cultured in non-differentiation medium. Likewise, the cell-seeded scaffolds cultured in differentiation medium exhibited a more intense red color upon staining with ARS, indicating greater mineralization compared to the blank scaffolds or the cell-seeded scaffolds cultured in non-differentiation medium. Background staining was observed in the blank scaffolds, potentially due to the presence of CaCl2 in the decellularization protocol.
Staining (H&E and VK) was performed on the scaffolds to analyze cell infiltration and mineralization, and SEM and EDS were used to further evaluate mineralization (Figure 2). H&E staining (Figure 2A) showed good cell infiltration in the cell-seeded scaffolds cultured in non-differentiation or differentiation medium. Multiple nuclei were visible in the periphery and throughout the scaffolds. The presence of collagen was also observed in the scaffolds in pale pink. In addition, VK staining performed on the scaffolds after 4 weeks of culture in differentiation medium revealed that the pore walls were stained whereas calcium deposits were solely detected along the outer edges of the pore walls in the scaffolds cultured in non-differentiation medium and may have resulted from calcium absorption during the decellularization treatment. Localized mineralization on the surface of the cell-seeded scaffolds cultured in differentiation medium for 4 weeks was observed by SEM analysis (Figure 2B). More specifically, mineral deposits resembling spheroid aggregates were observed on the periphery of the pores. In contrast, no mineral aggregates were observed on the blank scaffolds or the cell-seeded scaffolds cultured for 4 weeks in non-differentiation medium. Distinct characteristic peaks corresponding to phosphorous (P) and calcium (Ca) were observed in the EDS spectra of the selected regions of interest, specifically on the mineral deposits observed on the cell-seeded scaffolds cultured for 4 weeks in differentiation medium (Figure 2B).
In vitro biomechanical analysis (Figure 3)
The Young's modulus of the cell-seeded scaffolds was measured after 4 weeks of culture in either non-differentiation or differentiation medium (n = 3 for each experimental condition). It was compared to Young's modulus of the blank scaffolds (scaffolds without seeded cells) (Figure 3). No significant difference was observed in the modulus between the blank scaffolds (31.6 kPa ± 4.8 kPa) and the cell-seeded scaffolds cultured in non-differentiation medium (24.1 kPa ± 8.8 kPa; p = 0.88). In contrast, a significant difference was noted between the modulus of the blank scaffolds (31.6 kPa ± 4.8 kPa) and that of the cell-seeded scaffolds cultured in differentiation medium (192.0 kPa ± 16.6 kPa; p < 0.001). Additionally, a significant difference (p < 0.001) was observed between the Young's moduli of the cell-seeded scaffolds cultured in non-differentiation and differentiation media. Supplementary Figure 1 shows a typical stress-strain curve for the Young's modulus calculation.
In vivo biomechanical performance and bone regeneration (Figure 4 and Figure 5)
Surgical craniotomies were conducted on a total of 6 Sprague-Dawley rats. Bilateral 5-mm diameter defects were created in both parietal bones of the skull using a trephine bur, and apple-derived cellulose scaffolds without seeded cells were implanted in the calvarial defects (Figure 4A). After 8 weeks of implantation, the animals were euthanized, and the upper part of their skulls was collected and processed for either mechanical testing or histological analysis.
Based on visual assessment, the scaffolds seemed well integrated in the skull surrounding tissues. Mechanical push-out tests were performed to quantitatively assess the integration of the scaffolds (n = 7) in the host calvaria. The measurements were conducted using a uniaxial compression device (Figure 4B) immediately after the euthanasia of the animals. Results revealed that the peak force was 113.6 N ± 18.2 N (Table 1).
Histological analysis was conducted to assess cell infiltration and deposition of extracellular matrix within the implanted scaffolds (Figure 5). H&E staining revealed cellular infiltration within the scaffold pores and evidence of vascularization, as shown by the presence of blood vessels within the scaffolds. Additionally, MGT staining demonstrated the presence of collagen within the scaffolds.
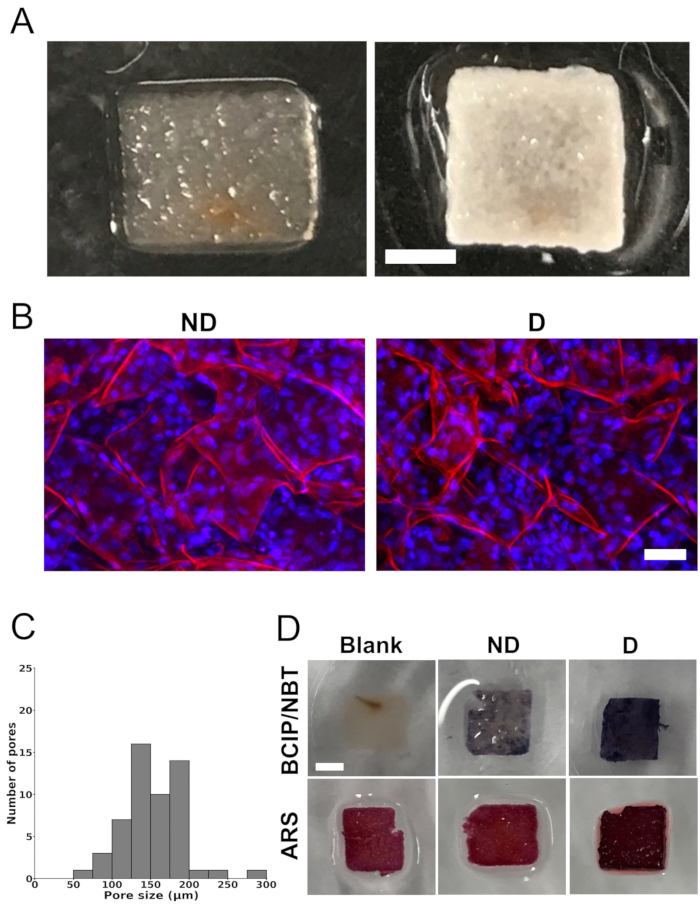
Figure 1: Scaffold images, pore-size distribution, and in vitro mineralization. (A) Representative photographs of an apple-derived cellulose scaffold after removal of the native cells and surfactant (left) and a scaffold seeded with MC3T3-E1 cells after 4 weeks of culture in osteogenic differentiation medium (right). Scale bar represents 2 mm. (B) Representative confocal laser scanning microscope images showing seeded cells in apple-derived cellulose scaffolds after 4 weeks of culture in non-differentiation medium ("ND") or osteogenic differentiation medium ("D"). Scale bar represents 50 µm. Staining was performed on the scaffolds for cellulose (red) using propidium iodide and for cell nuclei (blue) using DAPI. (C) Pore size distribution of decellularized apple-derived cellulose scaffolds, before being seeded with MC3T3-E1 cells, from maximum projections in the z-axis of confocal images. The analysis was performed on a total of 54 pores in 3 different scaffolds (6 pores in 3 randomly selected regions of interest per scaffold). (D) Representative images of scaffolds stained with 5-bromo-4-chloro-3'-indolyphosphate and nitro-blue tetrazolium (BCIP/NBT) to assess alkaline phosphatase (ALP) activity and with alizarin red S (ARS) to visualize calcium deposition, indicating mineralization (scale bar = 2 mm – applies to all). The scaffolds labeled as "blank" (scaffolds without seeded cells) showed no staining with BCIP/NBT, indicating the absence of ALP activity. On the other hand, the cell-seeded scaffolds cultured in differentiation medium ("D") displayed higher ALP activity, indicated by a more intense blue color, compared to the cell-seeded scaffolds cultured in non-differentiation medium ("ND"). For ARS staining, both the blank scaffolds and the scaffolds cultured in non-differentiation medium ("ND") exhibited a lighter shade of red compared to the scaffolds cultured in differentiation medium ("D"). The presence of calcium deposition in the scaffolds cultured in differentiation medium ("D") was illustrated by an intense deep red color. Each analysis was performed on three different scaffolds (n = 3). This figure was adapted with permission from Leblanc Latour (2023)27. Please click here to view a larger version of this figure.
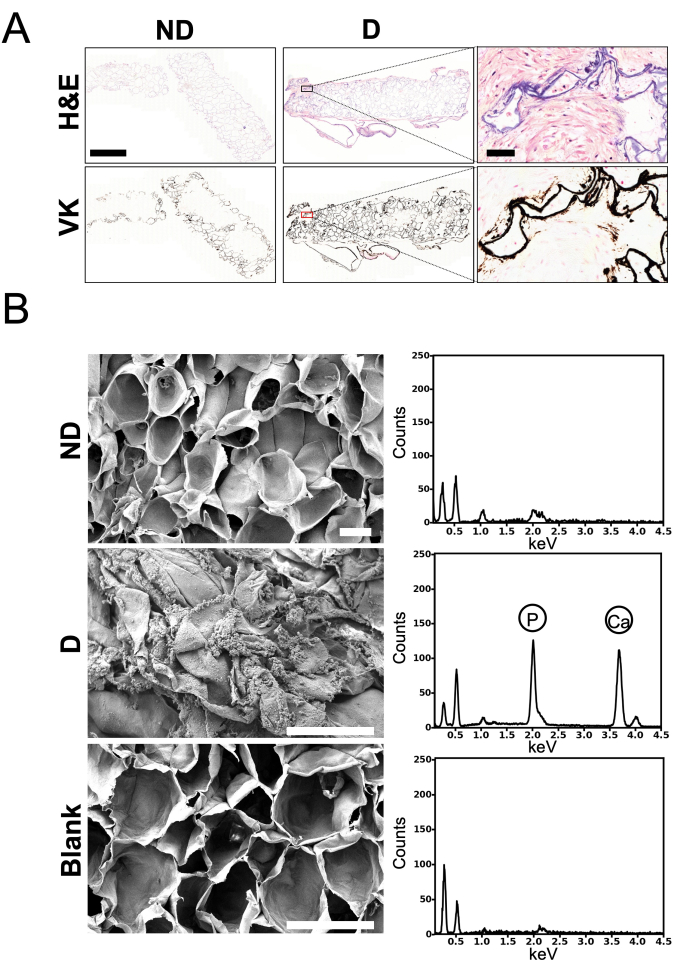
Figure 2: Histology, scanning electron microscopy (SEM), and energy-dispersive spectroscopy (EDS) analysis of in vitro scaffolds. (A) Representative images of top histological cross-sections of the scaffolds. Paraffin-embedded scaffolds were sliced into 5-µm thick sections that were stained with hematoxylin and eosin (H&E) to visualize cell infiltration, or with von Kossa (VK) to visualize mineralization within the scaffolds. Scaffolds were infiltrated with MC3T3-E1 cells, as shown by blue (nuclei) and pink (cytoplasm) staining visible at the periphery and throughout the scaffolds. Collagen (pale pink) was also visible (zoomed-in inset of "H&E – D"). Mineralization was observed only at the periphery of the pore walls in the scaffolds cultured in non-differentiation medium ("ND"). The pore walls in the scaffolds cultured in differentiation medium ("D") were entirely stained in black. The analysis was performed on one scaffold cultured in non-differentiation medium ("ND") and on two scaffolds cultured in differentiation medium ("D") (Scale bar for the lower magnification pictures = 1 mm, scale bar for the higher magnification pictures = 50 µm). (B) Representative micrographs obtained by SEM as well as EDS spectra. The scaffolds underwent sputter coating with gold and were imaged using a field-emission scanning electron microscope at a voltage of 3.0 kV (scale bar = 100 μm – applies to all). EDS spectra were acquired on each scaffold. Phosphorus (2.013 keV) and calcium (3.69 keV) peaks are denoted on each EDS spectrum. Both SEM and EDS were performed on three different scaffolds. Blank: scaffolds without seeded cells. This figure was adapted with permission from Leblanc Latour (2023)27. Please click here to view a larger version of this figure.
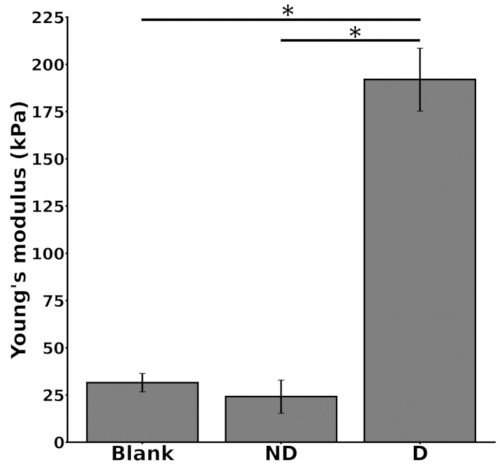
Figure 3: Young's moduli of in vitro scaffolds after 4 weeks of culture in either non-differentiation medium ("ND") or differentiation medium ("D"). Data are presented as mean ± standard error of the mean (SEM) of three replicate samples for each condition. Statistical significance (* indicates p<0.05) was determined using a one-way analysis of variance (ANOVA) and the Tukey post-hoc test. Blank: scaffolds without seeded cells. This figure was adapted with permission from Leblanc Latour (2023)27. Please click here to view a larger version of this figure.

Figure 4: Scaffold photograph prior to implantation and push-out test after 8 weeks of implantation: (A) Representative photograph of a scaffold prior to implantation; (B) Uniaxial compression device used for the push-out tests, with the load cell indicated by an asterisk (*) and the sample indicated by an arrow. This figure was adapted with permission from Leblanc Latour (2023)27. Please click here to view a larger version of this figure.
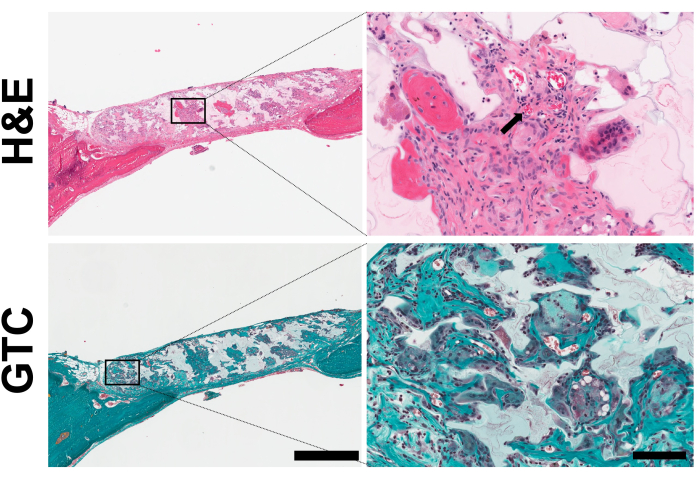
Figure 5: Histology analysis of in vitro scaffolds. Representative images of histological cross-sections from non-seeded scaffolds after 8 weeks of implantation. The sections were stained with either hematoxylin and eosin (H&E) to visualize cells or Masson-Goldner's trichrome (MGT) to visualize collagen. The arrow indicates red blood cells. The presence of collagen is visible (scale bar = 1 mm and 200 μm for the left and right insets, respectively). This figure was adapted with permission from Leblanc Latour (2023)27. Please click here to view a larger version of this figure.
| Sample number | Peak force (N) |
| 1 | 92.8 |
| 2 | 162.7 |
| 3 | 140.3 |
| 4 | 135.7 |
| 5 | 37.7 |
| 6 | 157.8 |
| 7 | 67.9 |
| Mean | 113.6 |
| SEM | 18.2 |
Table 1: Measured peak force from push-out tests.
Supplementary Figure 1: Typical stress-strain curve for Young's modulus calculation. Please click here to download this File.
