We performed the microdissection, staining, and imaging of the PVs in 10 12-16-week-old mice. Following the protocol, we successfully microdissected PVs together with the LA in all experimental mice and obtained sections with a comprehensive view of the PVs in eight mice. Overview images were taken at 10x magnification to identify the PV orifice (PVO) region at the LA-PV junction, the extrapulmonary PVs (PVex) (PVs in between the lung hilum and the LA-PV junction), and the intrapulmonary PVs (PVin) (PVs surrounded by lung tissue) (Figure 2). Zoomed-in images of the mentioned regions were obtained at a 40x magnification objective (Figure 3).
We found specific cTNT signals with a typical muscular striation in the PVO, the PVex, and the PVin (Figure 3). Cx43 was found in all three PV regions and was mostly projected between neighboring cardiomyocytes (Figure 3 and Figure 4A). Cx43-related signals were observed at both the polar side (Figure 3, yellow arrows) and the lateral side of the cardiomyocytes in the MS (Figure 3, red arrows).
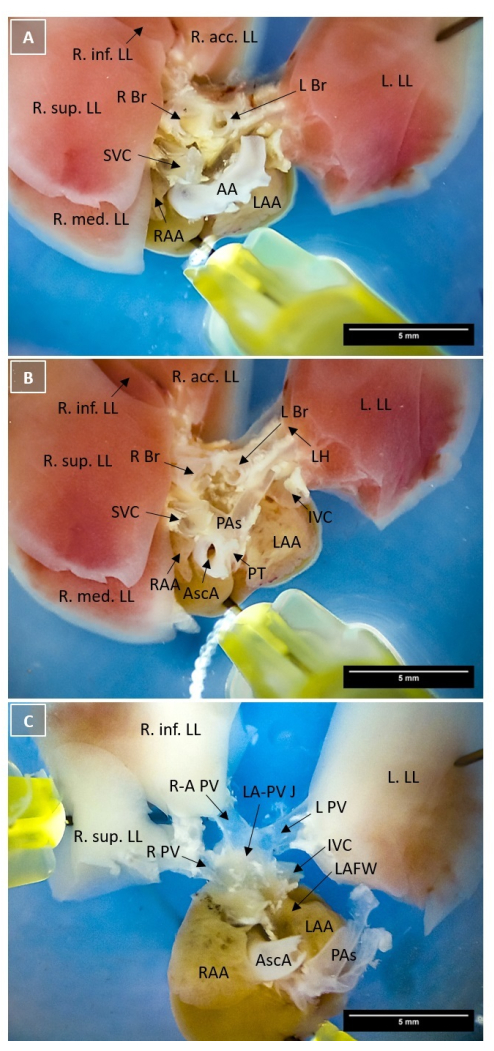
Figure 1: Identification of murine pulmonary veins under the microscope. Brightfield microscopy (10x magnification). The heart is pinned to the dissection dish with four pins at the atrium, the left lung lobe, the right middle lung lobe, and the right inferior lung lobe. (A) Overview of the heart base after separating the ventricular tissue from the atrium with the aortic arch in the middle. The atrium is located at the bottom; the lung lobes are spread out in the upper half of the picture. PVs are concealed by the AA, the main bronchi, and the PAs. (B) Top view image of the heart base after removing trachea, parts of the main bronchi, the aortic arch, as well as connective and fat tissue. The PAs in the middle of the image show their full course from the PT to the LH and cover the PVs. (C) Top view image of the PVs after cutting the PAs from the lung lobes and removing the extrapulmonary bronchial tissue. Scale bars = 5 mm. Abbreviations: AA = aortic arch; AscA = ascending aorta; IVC = inferior vena cava; L Br = left main bronchus; L PV = left pulmonary vein; LA-PV J = left atrium-pulmonary vein junction; LAA = left atrial appendage; LAFW = left atrial free wall; LH = lung hilum; L. LL = left lung lobe; PAs = pulmonary arteries; PT = pulmonary trunk; R Br = right main bronchus; R PV = right pulmonary vein; R-A PV = right ascending pulmonary vein; RAA = right atrial appendage; R. acc. LL = right accessory lung lobe; R. inf. LL = right inferior lung lobe; R. mid. LL = right middle lung lobe; R. sup. LL = right superior lung lobe; SVC = superior vena cava. Please click here to view a larger version of this figure.
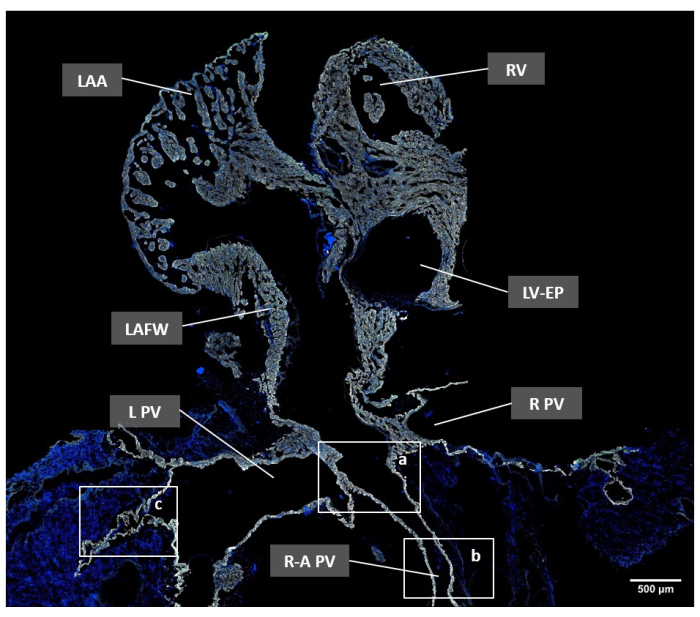
Figure 2: Overview of the adult mouse pulmonary veins and the left atrium in a transversal section image by immunofluorescence microscopy. Cell nuclei were stained with Hoechst-33342 (blue); cardiomyocytes were labeled with anti-cTNT antibody (white); and gap junctions were detected using anti-Cx43 antibody (green). The white rectangles highlight the following regions: a, PV orifice (PVO); b, extrapulmonary PV (PVex); and c, intrapulmonary PV (PVin). Scale bar = 500 µm. Abbreviations: L PV = left pulmonary vein; LAA = left atrial appendage; LAFW = left atrial free wall; LV-EP = left ventricle – ejection path; R PV = right pulmonary vein; R-A PV = right ascending pulmonary vein; RV = Right ventricle. Please click here to view a larger version of this figure.
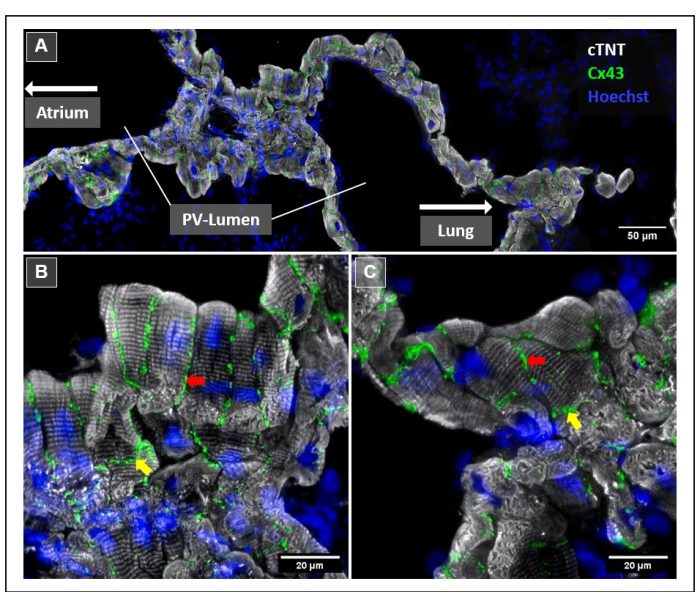
Figure 3: Zoomed-in view of the right PV myocardium using immunofluorescence microscopy. Cell nuclei were stained with Hoechst-33342 (blue); cardiomyocytes were labeled with anti-cTNT antibody (white); and Cx43+ gap junctions were detected by anti-Cx43 antibody (green). (A) EDF tile scans showing a longitudinal section through the right extrapulmonary PV. The LA and PV orifice are not shown in the image and would be located to the left, while intrapulmonary PV and corresponding lung parenchyma (also not shown) would be located to the right. (B,C) EDF single tile scans of the right extrapulmonary PV. Arrows highlight Cx43+ gap junctions at the cellular borders of cardiomyocytes. The yellow arrows indicate gap junctions at the polar sides of cardiomyocytes, colocalized with intercalated discs, while the red arrows indicate gap junctions at the lateral side of cardiomyocytes. Scale bars = 50 µm (A), 20 µm (B,C). Abbreviations: PV = pulmonary vein; EDF = extended depth of field; LA = left atrium. Please click here to view a larger version of this figure.
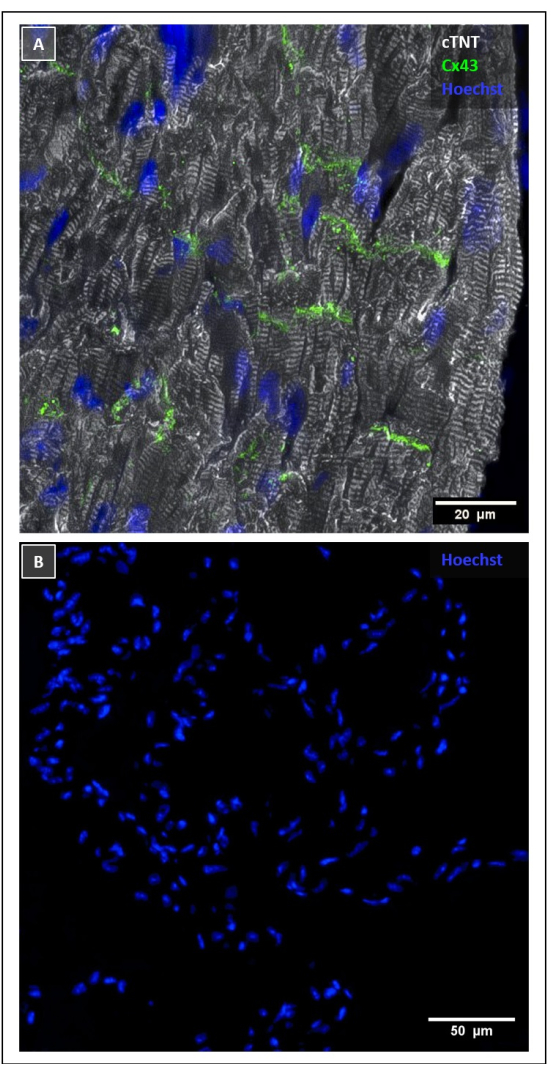
Figure 4: Immunofluorescence staining of control tissue. (A) Positive control staining of the murine LV. Cell nuclei were stained with Hoechst 33342 (blue); cardiomyocytes were labeled with anti-cTNT antibody (white); and Cx43+ gap junctions were detected with anti-Cx43 antibody (green). (B) Negative control staining of the right extrapulmonary PV by using secondary antibodies only. Cell nuclei are stained with Hoechst 33342 (blue). Scale bars = 20 µm (A), 50 µm (B). Abbreviations: LV = left ventricle; PV = pulmonary vein. Please click here to view a larger version of this figure.
| Filter-Cube | Excitation Filter | Emission Filter |
| DAP | 350 / 50 | 460 / 50 |
| L5 | 480 / 40 | 527 / 30 |
| TXR | 560 / 40 | 630 / 75 |
Table 1: Excitation and emission filter characteristics of the Leica DM6 B filter cubes.
| Compound | Final concentration | g or mL / 100 mL required |
| Fixing solution | ||
| Paraformaldehyde (PFA) 16% | 4% | 25 mL |
| Phosphate Buffered Saline (PBS) 1x concentrated | 75 mL | |
| Permeabilization Solution | ||
| Triton X-100 | 0.1% | 0.1 mL |
| Phosphate Buffered Saline (PBS) 1x concentrated | 99.9 mL | |
| Blocking Buffer | ||
| Normal goes serum (NGS) | 10% | 10 mL |
| Bovine Serum Albumin (BSA) | 0.5% | 0.5 mg |
| Tween 20 | 0.1% | 0.1 mL |
| Phosphate Buffered Saline (PBS) 1x concentrated | 89.9 mL | |
| Washing Buffer | ||
| Bovine Serum Albumin (BSA) | 0.5% | 0.5 mg |
| Tween 20 | 0.1 mL | |
| Phosphate Buffered Saline (PBS) 1x concentrated | 99.9 mL | |
Table 2: Buffer Recipes.
