Our embryo isolation procedure (Figure 1) allows isolation of up to 40 embryos in 4 hr if washes are performed, e.g. for molecular applications, or in less than an hour if washes are omitted, e.g. for cytological applications. Figure 2 displays high and low quality microcapillary tips and Figure 3 shows the setup of the embryo isolation machine. Figure 4 displays the process of embryo isolation on the inverted microscope.
We successfully applied our procedure for various applications published in several recent articles. The method was originally developed to analyze the parental contribution to the early embryonic transcriptome. Hybrid embryos generated through crosses between two different Arabidopsis accessions (Landsbergerecta (Ler) and Columbia (Col-0)) were isolated at the 2-4 cell stage. Total RNA was extracted, cDNA libraries were produced using linear amplification and sequenced on a SOLiD platform. The allele-specific transcriptomes were generated based on SNP analysis 8. To control our embryonic cDNA libraries prior to sequencing we amplified embryo-specific transcripts, WUSCHEL-HOMEOBOX 2, 8 and 9 (WOX2, WOX8, WOX9) and ACTIN 11 (13,14, Figure 5A).
Additionally, this method was used to isolate young embryos to analyze the embryonic DNA-methylation patterns at specific loci in the genome. Embryos were isolated and washed in 1x TE-buffer. Small-scale bisulfite sequencing was performed as described 9,15.
Similarly, embryo isolation has proved very useful in reporter gene assays with low embryonic expression, and where the maternal seed coat confounds detection or is masked by reporter expression in tissues surrounding the embryo (endosperm, seed coat). Embryos carrying the reporter transgene are stained (ß-glucuronidase; GUS) or directly analyzed (Green or Red Florescent Protein; GFP, RFP) following isolation. An example is given in Figure 5B for embryos expressing a GUS reporter gene under the control of the MEDEA promoter (pMEA) 10. The relatively ease with which many embryos can be isolated also allows for quantitative analyses (e.g. number of embryos stained in different genetic backgrounds, Raissig, Grossniklaus et al. in preparation).
Finally, we successfully applied Fluorescent In Situ Hybridization (FISH) and immunostaining techniques to study the nuclear architecture in isolated embryos embedded in acrylamide pads on slides. An example of FISH using probes against the centromere repeats and nucleolar organizing regions is shown in Figure 5C.
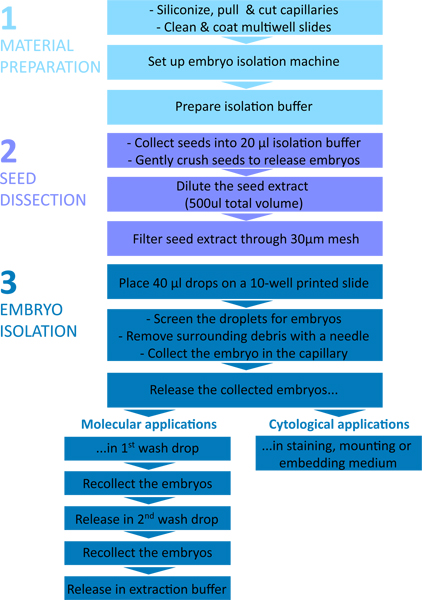
Figure 1. Flow chart of the embryo isolation procedure. The protocol is divided in three parts: 1 – Material preparation; 2 – Seed dissection; 3 – Embryo isolation.

Figure 2. Microcapillary tips. A. High-quality microcapillary tips with a smooth opening of around 100 μm. B. Low-quality microcapillary tips with wider opening and irregular contour (those tips are acceptable for cytological applications only). Scale bar: 2 mm.
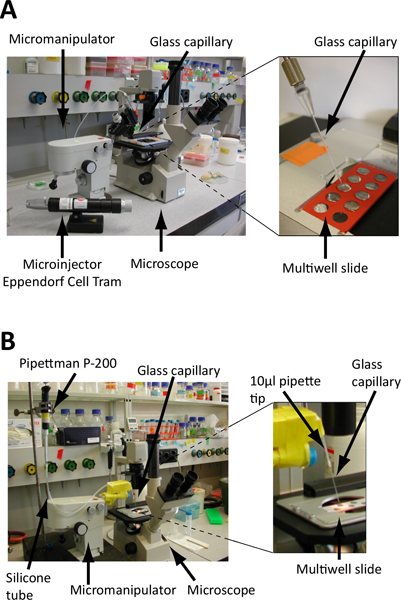
Figure 3. Set up of the embryo isolation microscope. A. and B. Screening is performed on microscope slides under an inverted microscope. Embryos are collected using a glass microcapillary fixed on a micromanipulator to precisely control its position (see also Figure 4) and connected to either a microinjector (A) or standard laboratory pipette (B). A. The glass microcapillary is linked to a hydraulically controlled microinjector (e.g. Eppendorf Cell Tram Vario) for a precise control during embryo collection B. The glass microcapillary is attached to a standard P-200 pipette via a rubber flex tube. Embryo collection and release is controlled by turning the calibration wheel of the pipette. Cut-end pipette tips are used as connectors and the junctions are sealed with parafilm. The microcapillary is fixed on the micromanipulator via polystyrene blocks, Falcon tubes and tape.
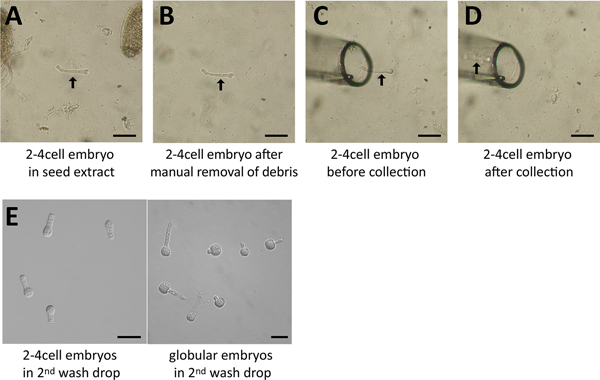
Figure 4. Embryo isolation process. A. A 2-4 cell embryo (arrow) was identified in the seed extract (screening droplet) and is surrounded by debris (seed coat fragments). B. The surrounding debris was removed manually using a needle. C. The glass capillary was moved right beside the embryo (arrow). D. The embryo was collected and is now within the capillary (arrow). E. Several embryos in the last drop following washes. Scale = 50 μm.
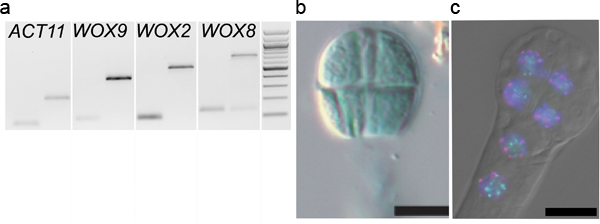
Figure 5. Downstream applications of embryo isolation. A. Gene expression analyses: PCR amplification of WOX9, WOX2, WOX8 and ACTIN11 13,14 on cDNA libraries (generated as described 8) from 2-4 cell embryos washed 1x times (1st lane) and on genomic DNA (2nd lane). B. Reporter assay: An 8-cell embryo isolated from plants carrying a pMEA::GUS construct was stained on a slide in a standard GUS staining solution (reproduced from Raissig et al., 2011 after permission from The Plant Cell, ASPB copyright). C. Fluorescent In-Situ Hybridization (FISH): an 8-cell embryo was hybridized with probes against centromeric repeats (red), and 45S rDNA repeats (green) before indirect immunodetection 16. The dual-color FISH images were overlaid with the DAPI counterstaining and DIC images (DM6000B epifluorescence microscope, Leica, Germany).
| Application | Isolation buffer | Destination buffer |
| RNA extraction (e.g. for amplification and transcriptomics) | 1x First-Strand buffer (Invitrogen), 1 mM DTT, 4% RNAseOUT | RNA extraction buffer |
| DNA extraction (e.g. for bisulfite sequencing) | 100 mM Tris, 10 mM EDTA, pH 8 (TE) | DNA extraction buffer or 1x TE buffer |
| GFP reporter analysis | 1 M Glycin in 0.5x MS1 | 1 M Glycin in 0.5x MS |
| GUS reporter analysis | staining buffer2 without X-Gluc (substrate) | staining buffer with X-Gluc (substrate) |
| FISH & Immunostaining | 100 mM Phosphate Buffered Saline (PBS) | activated Acryl:Bisacryl mix (33:3) on a Superfrost Plus slide -polymerize for 30 min |
Table 1. Isolation and destination buffers for different downstream applications. The buffers can be adapted to specific experimental requirements. 1 Murashig & Skoog medium. 2 e.g. Jefferson et al., EMBO J. 6 (13), 3901-3907 (1987).
