Den mekaniske mikromiljøet spiller en viktig rolle i mange cellefunksjoner, som proliferasjon, migrering og differensiering, som har en betydelig innflytelse på utvikling og homeostase av vev, og også i sykdommer 1-6. Gjennom årene har en rekke eksperimentelle verktøy blitt brukt til mekanisk stimulere celler eller vev og måle mekaniske egenskapene til biologiske vev med mål om å øke vår forståelse av grunnleggende mechanobiology og studere utbruddet og progresjon av sykdommer 6-17. Imidlertid må man ofte avhengige av flere ulike eksperimentelle enhetene for å oppnå målene i en bestemt studie. Denne artikkelen presenterer et enkelt multi-funksjonelle, dual-aksen strekker (BAXS) plattform, som gjør det mulig for studier som undersøker den rollen som mekaniske egenskaper og mekaniske krefter spille i biologi ved sub-mobilnettet til hele vev lengdeskalaer. Den BAXS plattformen ikke bare åpner for quantification av de mekaniske egenskapene til isolerte vev, men også forenkler muligheten til å bruke enkle, komplekse og dynamiske belastnings felt til levende celler for å forstå sine svar til strekking som skjer in vivo. Den BAXS plattformen opprettholder også kapasitet til å utføre live-cell mikros under mekanisk testing og forstyrrelser på celler og vev.
Den BAXS plattformen er en spesialbygd anordning som kan brukes for å undersøke effekten av substratet deformasjon på cellenivå, og å utføre strekk-tester av biologiske vev (Figur 1a). En aluminiumvarmeanordning ble fremstilt for å få plass til en standard 10 cm petriskål og opprettholde noen fysiologiske løsninger ved 37 ° C ved hjelp av en temperaturregulator og Kapton beredere (figur 1B). Denne BAXS plattformen kan integreres på en omvendt fasekontrast-og / eller fluorescens mikroskop og muliggjør simultan avbildning (figur 1C).I korte trekk består BAXS plattform av fire lineære talespole motorer hvorav de bevegelige delene er montert på miniatyr lineær bevegelse kulelager lysbilder orientert langs to vinkelrette akser (figur 1D). En lineær posisjonering stadium er montert på hver av de fire motorer for å tillate vertikal bevegelse av klemsystemet som skal brukes (figur 1E). Posisjonen til hver motor er overvåket av en optisk giver med en oppløsning på 500 nm (fig. 1F). Alle fire motorene styres uavhengig av hverandre med en bevegelseskontroller ansette optisk encoder tilbakemelding til å utføre bevegelseskommandoer (figur 1G). En LabVIEW grensesnitt gir full kontroll over omfanget forskyvning, hastighet og akselerasjon for hver motor for å generere helt tilpasses, statisk og dynamisk, deformasjon av cellene eller vevsprøver.
Den teknikk som brukes for å fremkalle en deformasjon i cellene er oppnådd ved ganske enkelt å allowing celler for å feste seg til en fleksibel og gjennomsiktig substrat, og deretter strekking av dette substrat ved hjelp av de fire motorene i BAXS plattformen. Den BAXS plattformen tillater montering av noen spesialdesignede sett med klemmer for å feste underlaget på talespole motorer. For dette formål utformet vi et sett av klemmer til hvilke en fleksibel og gjennomsiktig substrat, som består av polydimetylsiloksan (PDMS), kan festes (figurene 2A-C og figur 3). Ettersom klemmene vil bli utsatt for fysiologiske løsninger, alle deler maskinert av rustfritt stål for å gi rom for sterilisering. Disse klemmene har blitt nøye designet for å bringe underlaget så nært som mulig til mikroskop mål å forbedre bildekvaliteten, samtidig som belastningen på underlaget under stretching (figur 2D).
Den samme BAXS plattformen kan også brukes til å kvantifisere stivhet i små vevsprøver, med et passende sett av klemmer med tilpasted støtter for vevsprøver og en lastcelle for å overvåke krefter. Flere tilnærminger kan tas for å montere en vev til BAXS plattform motorer; i dette tilfelle rustfrie insekt minutiens nålene kan hekte gjennom åpningen i vaskulære vev for å utføre strekk-tester (Fig. 4A-B). Alternativt, for tykke vev uten en naturlig åpning, vev kanter kan enten holdes i posisjon med klemmene festet til talespole motorer eller limt til små glassplater med biologisk lim og festet til motorene med klemmene. For å utføre strekk-tester en miniatyr-lastcelle er nødvendig, og kan lett tas bort på BAXS plattformmotorer og benyttes for å måle kraft som virker på vevet under et strekk syklus (figur 4C). Som BAXS plattformen er sammensatt av fire motorer, innføring av en andre lastcelle gjør det mulig å utføre strekk-testing langs to ortogonale retninger. Denne evnen gjør det mulig å quantify den mekaniske stivhet av en enkelt vev langs to vinkelrette retninger i samme eksperiment.
Viktigere, i alle konfigurasjoner, cellene eller vevsprøver av interesse blir alltid opprettholdt i et temperaturkontrollert bad som er tilgjengelig for brukeren. Denne evnen gjør det mulig for innføring av farmakologiske midler i prøven som strekker seg for å undersøke tidsmessige reaksjon av prøven. I tillegg, som den optiske aksen til det inverterte mikroskop forblir uforstyrret, alle former for mikroskopi er fremdeles tilgjengelig for brukeren. Til slutt, som alle fire motorene i BAXS plattformen er uavhengige, er det mulig å anvende svært konfigurerbar belastningsfelt til prøven av interesse. In vivo-celler og vev er utsatt for komplekse og anisotrop strekking som kan være mer hensiktsmessig etterlignet i denne plattformen i motsetning til tradisjonell uniaxial strekker plattform 7,13,15,18,19. Dessuten, de fysiske egenskapeneav belastningen feltet kan endres på fly under et eksperiment. Disse egenskaper tillater at brukeren kan undersøke cellulær-og vevsnivå respons på et stort antall svært komplekse, anisotropisk, er tidsmessig og rommessig varierende belastningsfelt. Denne artikkelen beskriver de fordeler og begrensninger av BAXS plattform samt dens design, operasjonelle prinsipper, og de eksperimentelle detaljer for enkelt celle og hele vev eksperimenter.

Figur 1. Oversikt over BAXS plattformen. A) toppriss av BAXS plattform som viser de fire talespolemotorer. B) detaljert bilde av petriskålen varmeapparatet benyttes til å opprettholde celler og vev ved 37 ° C. C) Plattformen kan monteres på en invertert mikroskop for å utføre levende cell imaging under strekker eksperimenter.D) Detaljert bilde av talespolen motor; den bevegelige del av plattformen. E) detaljert bilde av den lineære posisjonstrinn som tillater vertikal forskyvning av klemmesystemer. F) detaljert bilde av den optiske giver som gir sanntid posisjonen av motoren til bevegelsesregulatoren. G) Detaljert bilde av bevegelseskontrolleren som viser de fire optiske enkoderinnganger og effekter til de fire talespole motorer.
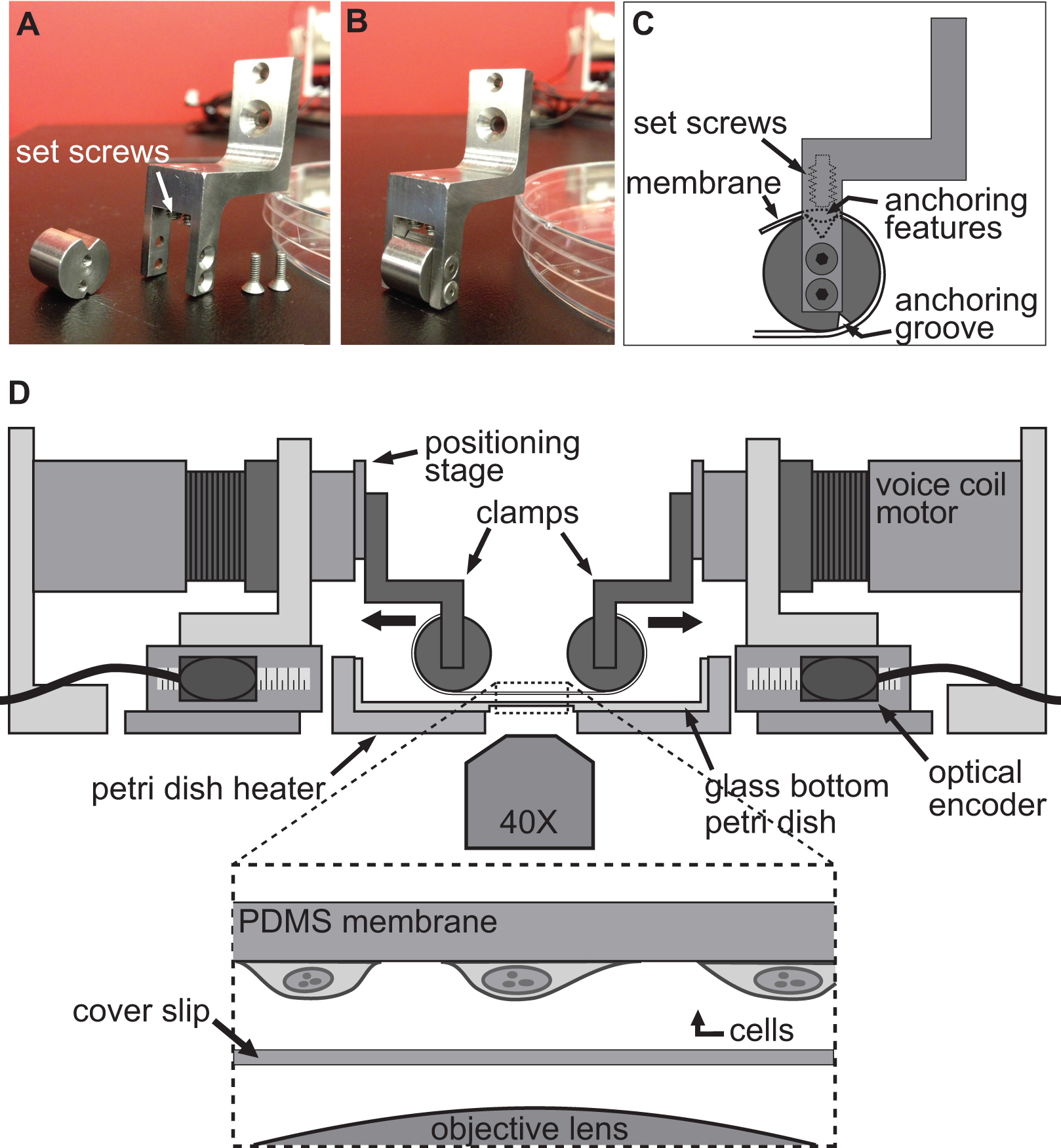
Figur 2. Klem system for celle strekker eksperimenter. AB) i bilder som viser detaljene av klemmer som brukes til å feste PDMS substrat til talespole-motorer for strekking. C) Substratet blir viklet rundt den sylindriske del av klemmen med dens forankring features sitter i sporet på toppen. Da underlaget er sikret ved hjelp av justeringsskruene som presser underlaget / forankring funksjoner inn i topp groove. D) Illustrasjon av BAXS plattform med klemmer som holder underlaget på plass. Det innfelte viser en detaljert visning av underlaget med celler som er knyttet til det sittende rett over et dekkglass og mikroskop mål.
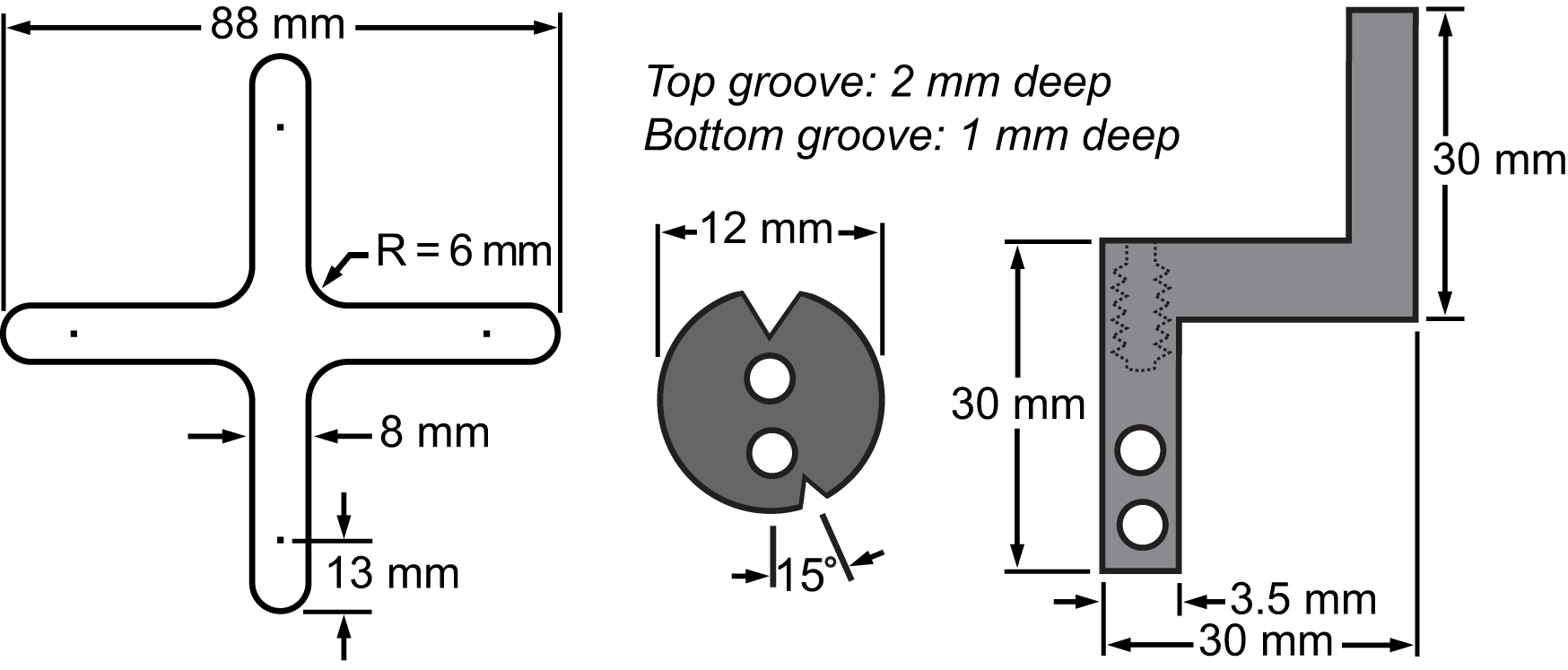
Fig. 3. Bill av materialer av membranen og dens klemsystemet. Tegninger som viser dimensjonene av de viktigste delene er integrert på biaksial plattformen for å utføre celle strekker eksperimenter.
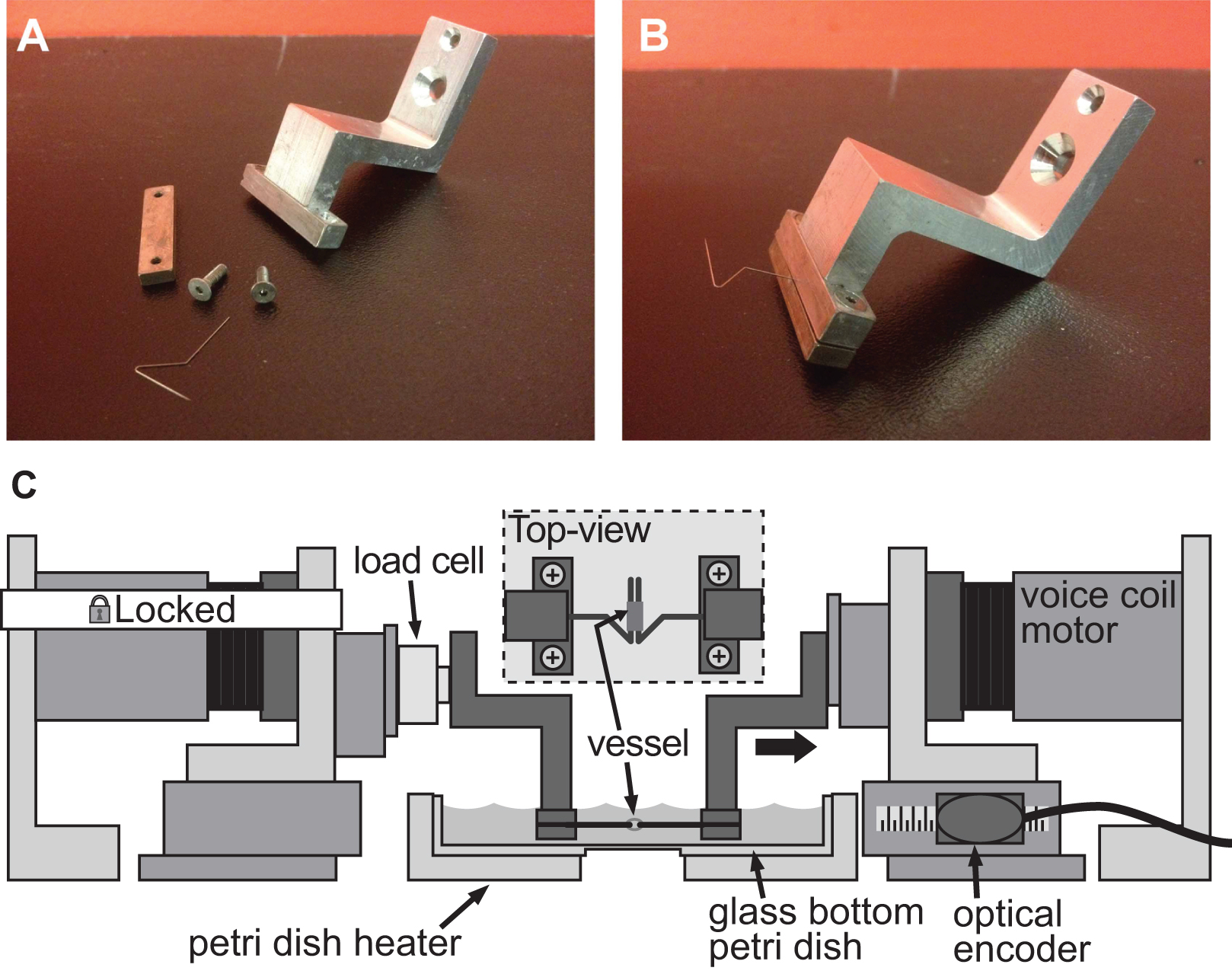
Figur 4. Exrikelig av et klemsystem for stivhet vurdering av småkalibrede fartøy. AB) detaljerte bilder av klemsystemet brukt til å indusere deformasjon i en 1 mm diameter mus aorta. Rustfritt stål nålene ble omhyggelig formet til åpne trekanter for å tillate fartøyet å gli på begge pinnene. C) Illustrasjon av BAXS plattform med klemmene som holder fartøyet og en lastcelle koblet mellom den faste motor, og den venstre klemme. Det innfelte viser et detaljert topp visning av montert på pinnene fartøy.
