קביעת כמותית של תגובות ביוכימיות מספקת תובנות לגבי התהליכים הביולוגיים העומדת בבסיס של חיים. Calorimetry מציע מתודולוגיה ללא תווית לאפיין כמעט בכל תגובה כימית בפתרון כמותית. טכניקה זו מודדת את החום שמשתחרר או נספג לאורך זמן, ולכן מערכת זיהוי אוניברסלית ומתודולוגיה נוחה מאוד לכמת את כמות המולקולות מגיבים (כלומר תרמודינמיקה מחייבת), כמו גם על מנת למדוד את קצב התגובה (כלומר קינטיקה). בפרט, calorimetry טיטרציה בידוד התרמי (ITC) כבר אימץ כשיטת בחירה לאפיין את התרמודינאמיקה של שיווי משקל biomolecular, הכולל חלבון ליגנד, חלבונים, יוני חלבון מתכת ואינטראקציות החלבון-DNA 1-6. בנוסף, היכולת של ITC לספק מידע הקינטית הופכת אותו למערכת חזקה מאוד למדוד קטליזה אנזים, למרות שהפוטנציאל של יישום זה הוא עדייןלזלזל 7-9.
משוואת מיכאליס מנטן-10 היא תיאור כמותי של תגובות אנזימטיות, כפי שהוא מספק קשר בין קצב התגובה וריכוז המצע, תלוי בשני פרמטרים הקינטית: קבוע מיכאליס (K M) ושיעור קבוע קטליטי (k חתול) . K החתול / יחס K M הוא כאמור את היעילות הקטליטית של אנזים. בפועל, קביעת K M ו-k חתול לתגובה ספציפית מספק תיאור של קטליזה מלאה.
בתגובה האנזימטית טיפוסית (איור 1), מצע (S) אינטראקציה עם האנזים (E) ויוצר מורכבות אנזים-מצע (ES), שמופעל בהמשך למצב המעבר (ES *). זה האחרון הופך למורכב אנזים תוצר הלוואי (EP) שסופו של הדבר dissociates. צעד הבאיםים מתוארים על ידי התגובה הבאה.
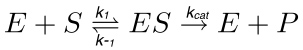 (1)
(1)
כאשר k 1 הוא השיעור קבוע להקמת קומפלקס ES, k -1 הוא שיעור קבוע לדיסוציאציה של קומפלקס ES, בעוד k החתול הוא שיעור קבוע קטליטי או מספר מחזור.
לפי משוואת מיכאליס מנטן-10, שיעור התגובה יכול להיות מחושב כ:
 (2)
(2)
שבו חתול K = M (k -1 + K חתול) / k 1 ו-k = מקסימום v / [E], עם מקסימום v להיות המהירות המקסימלי הגיעה כאשר כל האנזים קשור למצע.
Calorimeter טיטרציה הבידוד התרמית היא הכלי המשמש במחקר זה כדי לאפיין את הידרוליזה אנזימטית של אוריאה. מכשיר זה עשוי ממגן adiabatic המכיל שני תאים בצורת שטבע (איור 1). אלה מחוברים לחלק החיצוני עם צינורות גישה צרים. התא המדגם (בערך 1.4 מיליליטר) טעון עם פתרון האנזים, ואילו תא ההתייחסות הוא בדרך כלל מלא במים או בממסים המשמשים לניתוח. מזרק מסתובב עם מחט ארוכה והנעת מערבבים המצורפות, המכיל בדרך כלל CA. 0.3 מיליליטר של תמיסת מצע, הוא רכוב על התא המדגם. מכשיר תרמואלקטרי מודד את ההבדל בטמפרטורה בין המדגם ותא ההתייחסות ו, באמצעות "רשת משוב סלולרי", הוא שומר על ההבדל הזה באפס על ידי הוספה או הפחתת חום. במהלך הניסוי, המצע מוזרק לתוך תמיסת האנזים בטמפרטורה שנבחרה קבועה. כאשר enתגובת zymatic מתרחשת, כמות החום שמשתחררת או נספגת היא פרופורציונלית למספר מולקולות מצע, שהומרו למולקולות מוצר. בנוסף, שיעור זרימת חום קשור באופן ישיר לשיעור של התגובה. נתונים שנמדדו, המופיע כסטייה של עקבות חום מנקודת ההתחלה ראשונית (איור 1), מייצגים את כוח התרמיות (μcal / sec) המסופק על ידי המכשיר הסלולרי המדגם, שהוא פרופורציונאלי לזרימת החום המתרחשת בתא המדגם לאורך זמן.

איור 1. ייצוג סכמטי של calorimeter טיטרציה בידוד התרמית ללמוד תגובות אנזימטיות. תגובה האנזימטית מתרחשת על טיטרציה של המצע (במזרק להזרקה) לתוך תמיסת האנזים (בתא המדגם) גורם לשינוי של יסכוח mal שפורסם על ידי calorimeter, דרוש כדי לשמור על הפער של טמפרטורה בין תא דגימה וקבוע תא התייחסות. לחץ כאן לצפייה בתמונה גדולה יותר.
בסך הכל, השינוי בחום (Q) הוא פרופורציונאלי לאנתלפיה הטוחנות של התגובה (ΔH) ומספר השומות של מוצר שנוצר (n), אשר בתורו ניתן על ידי בסך הכל הפעמים נפח הריכוז:
 (3)
(3)
היווצרות המוצר לאורך זמן (DP / DT), אשר תואם את קצב התגובה, ולכן יכול להיות קשור לכמות החום שנוצר על אותו הזמן (DQ / dt) דרך היחס:
 (4)
(4)
על פי משוואה זו, על מנת לקבל מיכאליס-מנטן עלילה יש צורך למדוד i) סך ΔH אנתלפיה טוחנת, וii) את זרימת DQ / dt החום בריכוזים מצע שונים. בדרך כלל, זה מתבצע בשני ניסויים שונים: בניסוי הראשון (שיטת 1, M1), המצע מוזרק לתוך תמיסת האנזים והחום לגיור מצע שלם נמדד; בניסוי השני (השיטה 2, M2), זריקות מרובות של מצע מבוצעות וקצב ייצור חום נמדד בריכוזים מצע שונים. שתי מערכות אלו של נתונים מספיקות כדי לגזור את הפרמטרים הקינטית K M ו-k חתול.
במאמר הנוכחי, פרוטוקול כללי כדי לקבוע את הפרמטרים הקינטית לתגובות אנזימטיות בוצעו באמצעות ITC מתואר. אנחנו יישמנו את השיטה להידרוליזה אוריאה על ידי האוריאה ensiformis Canavaliase, כמערכת התייחסות. ההסכם הטוב בין התוצאות שהתקבלו בשיטה זו ונתונים שדווחו בספרות מדגים את האמינות של גישה זו.
