Luminescence Resonance Energy Transfer (LRET) er et derivat av den velkjente Fluorescence Resonance Energy Transfer (FRET) teknikk en. I likhet med slite, kan LRET brukes til å måle avstander og avstandsendringer mellom donor og akseptor fluoroforene knyttet til bestemte steder på protein av interesse innenfor intervallet 10-100 A 1-3. Prinsippene for LRET er også lik FRET ved at resonans energioverføring skjer mellom to fluoroforer proksimale når emisjonsspekteret av donor fluoroforen overlapper med absorbsjons-spekteret av akseptor fluoroforen. Effektiviteten av denne overføringen er relatert til avstanden mellom de to fluoroforer etter følgende ligning:
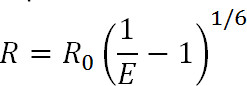 Ekv. 1
Ekv. 1
hvor R er avstanden mellom de to fluoroforer, E er effektiviteten av noErgy overføring, og R 0, omtalt nedenfor, er Förster radius for fluoroforen par, dvs. avstanden som effektiviteten i overføringen er halv maksimal. Fra denne ligning kan man se at effektiviteten er relatert til størrelsen av avstanden opphøyd i inverse sjette strøm 1. Disse inverse sjette strømavhengighet som gir mulighet for FRET og LRET målinger for å være utsøkt følsom selv for små avstandsendringer når nær R 0 av FRET paret. Evnen til å spesifikt merke ønskede steder på proteiner eller andre makromolekyler som tillater en å dra fordel av denne følsomhet til å overvåke konformasjonsendringer.
Sammenlignet med slite, som bruker konvensjonelle organiske fargestoffer, og tilbyr LRET ytterligere fordeler. I LRET, i stedet for ved hjelp av et organisk fargestoff som donor fluorofor, et lantanid-serien kation, typisk Tb 3 + eller 3 + Eu, anvendes 1,4-6. Fluoroforene som faller under denne kategori, f.eks, terbium chelat, er også meget anvendelig ved at de kan brukes sammen med et bredt spekter av akseptor fluoroforer. Denne fleksibiliteten er gjort mulig fordi emisjonsspektrene av chelaterte lanthanider inneholde flere skarpe utslipps topper, slik at for en enkelt art av donor fluoroforen til å brukes sammen med en av et bredt utvalg av akseptor fluoroforer. Dermed kan sensibilisert akseptor utslipp oppdages uten engstelse for å smitte gjennomslag fra donor utslipp fem. Den experimenter velger den spesifikke akseptor basert på den forventede avstanden mellom de to fluoroforer (figur 1 og tabell 1). I disse chelaterte lantanid fluoroforer, blir metallionet chelatert med et molekyl som inneholder en antennegruppe som sensitizes normalt dårlig absorberende lantanid til eksitasjon, så vel som en funksjonell gruppe bioreactive å tjore ion til en bestemt funksjonell gruppe på makromolekylet 1, 5,6. ONCe spent, lanthanider slappe av til grunntilstanden via utgivelsen av fotoner med en dempefaktor i millisekund området. Fordi forfallet er verken en singlet-til-sing avslapping eller en triplett-til-sing avslapning, utslipp av fotoner kan ikke riktig kalles fluorescens eller morilden, men er mer riktig betegnes luminescens en. Den lange nedbrytning av lanthanid luminescence bidrar sterkt i levetid målinger. Levetid målinger kan så brukes for å bestemme effektiviteten via følgende relasjon:
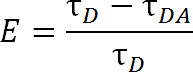 Ekv. 2
Ekv. 2
der, er E effektiviteten i overføringen, er τ D levetiden til donor (kjelatert lanthanid) når de ikke deltar i energioverføring, og τ DA er levetiden til donor når du deltar i energioverføring med akseptor. Med LRET, τ DA kan alså måles som levetiden på lysfølsomt akseptor utslipp fordi terbium levetid er så mye større enn en organisk akseptor fluoroforen. Den akseptor avgir med den samme levetid som dens eggende eksitasjon (donor lantanid), og noe bidrag til levetid fra akseptor egen iboende fluorescens levetid er forholdsvis ubetydelig. Ved å måle den sensibiliserte utslipp fremfor donor utslipp vi også eliminere behovet for å sikre at merkingen på nøyaktig en 1: 1-forhold av donor til akseptor. Protein kan i stedet merkes samtidig med både akseptor og donor fluorophores. Et heterogent merket befolkningen vil føre, men dobbel-donor merkede proteiner vil ikke slippe ut i akseptor bølgelengde og dobbelt akseptor merkede proteiner vil ikke være begeistret. Videre bør avstanden mellom fluoroforer være den samme, uavhengig av hvilken cystein side en gitt fluoroforen er festet til, spesielt ved bruk av den isotrope lanthanider som en donor, så need til å angi et gitt område for å motta enten donor eller akseptor er unødvendig. Intensitet kan påvirkes med en heterogen populasjon, men bør likevel være mer enn tilstrekkelig til å bli detektert.
Ved planlegging av eksperimenter, bør valget av fluoroforer være diktert av R-0-verdi av paret, så vel som de forventede avstandsområde som skal måles. R 0-verdien er definert ved den følgende ligning:
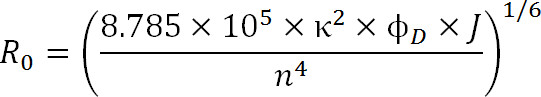 Ekv. 3
Ekv. 3
hvor, R 0 er Förster radius i Ångstrøm, κ 2 er orienteringen faktoren mellom de to fargestoffer (vanligvis antas å være 2/3), φ D er kvanteutbytte av donor, J er den spektrale overlapping integrert mellom donor emisjonsspektrum og akseptor er Absorpsjonsspekteret i M – 1cm -1 nm til 4, og n er brytningsindeksen for mediet 1.
Våre laboratorie har lagt en modifikasjon av den konvensjonelle LRET teknikken ved å innføre en protease-gjenkjenningssete mellom donor og akseptor etikett steder på proteinet som blir undersøkt. Denne modifikasjon gjør det mulig for undersøkelse i ikke-renset systemer som hele pattedyrceller 7. Denne teknikken er spesielt nyttig ved bruk av cysteiner som seter for merking, siden i ferd med merking med maleimide-konjugert fargestoffer som binder seg til cystein-sulfhydrylgrupper, andre proteiner i celler som har cysteiner er også merket. Imidlertid, ved å inkludere proteaseinhibitorer spaltningsseter for proteinet av interesse, og å måle levetider før og etter spalting, experimenter kan kvantitativt subtrahere bakgrunnssignalet etter protease-spalting fra rå signal. Denne subtraksjon isolerer det spesifikke signal som oppstår fra for proteinet av interesse (Figure 2). Ved hjelp av den modifikasjon som er beskrevet ovenfor, kan LRET brukes til å måle avstandsendringen mellom terbium chelat-donor og akseptor sonde på et protein og således overvåke konformasjonsendringer i proteinet, i nærheten av fysiologisk tilstand uten behov for rensing.
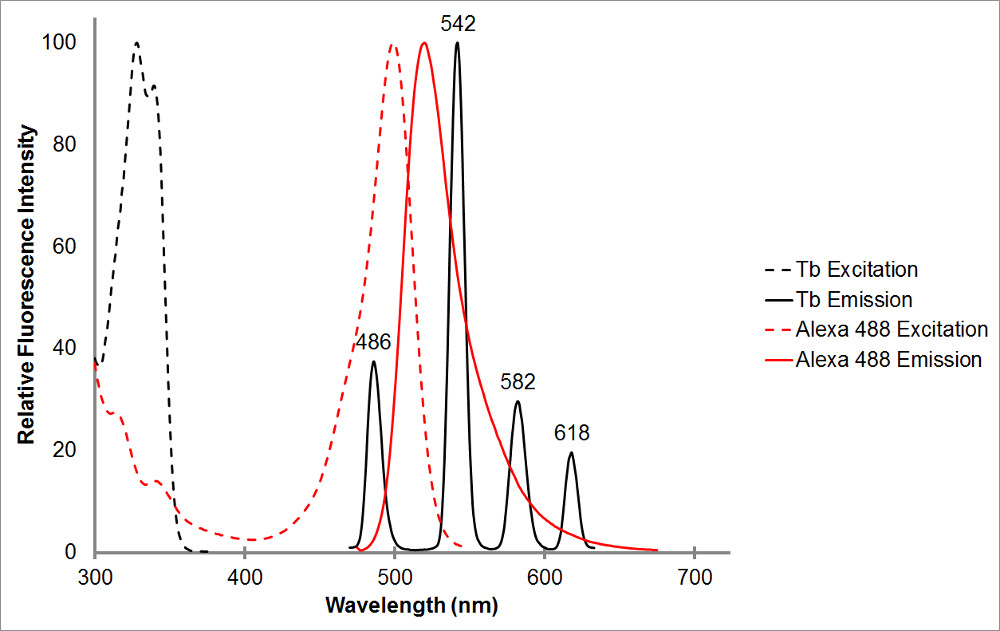
Figur 1. Den absorpsjon og emisjon spektra av chelatert terbium i svart, så vel som en representativ akseptor, Alexa 488, i rødt. Legg merke til de flere utslipps topper og den skarpe, smale emisjonsområde for hver topp av terbium chelat. Dette mønster gjør det mulig for terbium til å brukes sammen med en rekke akseptor fluoroforer og letter måling av lysfølsomt emisjon innenfor avstander hvor terbium viser ingen emisjon. Terbium utslipp topp på 486 nm lapper ganske godt med en bsorption toppen av Alexa 488, slik at for resonans energioverføring å oppstå mellom de to fluoroforer. En bølgelengde på 515 nm er et utmerket valg for å oppdage lysfølsomt utslipp for dette paret som det er i dalen mellom Terbium utslipps toppene, og ganske nær Alexa 488 utslipp topp på 520 nm. Legg merke til at det å være nær akseptor topp, selv om ønskelig, ikke er nødvendig-565 nm er fortsatt i stand til å oppdage Alexa 488 utslipp uten også å oppdage terbium utslipp.
| Akseptant fluoroforen | R 0 (Å) | Utslipp bølgelengde (nm) |
| Atto 465 | 36 | 3px; "> 508|
| Fluorescein | 45 | 515 |
| Alexa 488 | 46 | 515 |
| Alexa 680 | 52 | 700 |
| Alexa 594 | 53 | 630 |
| Alexa 555 | 65 | 565 | Cy3 | 65 | 575 |
Tabell 1. En liste over ofte brukte akseptor fluoroforene for LRET bruker terbium chelatet som donor 11. De R-0-verdiene ble målt når donor og akseptor ble festet til den oppløselige agonist bindende domenet av AMPA-reseptorer. Det er ideelt å måle R 0 verdien på nytt for hver nye systemet som studeres.
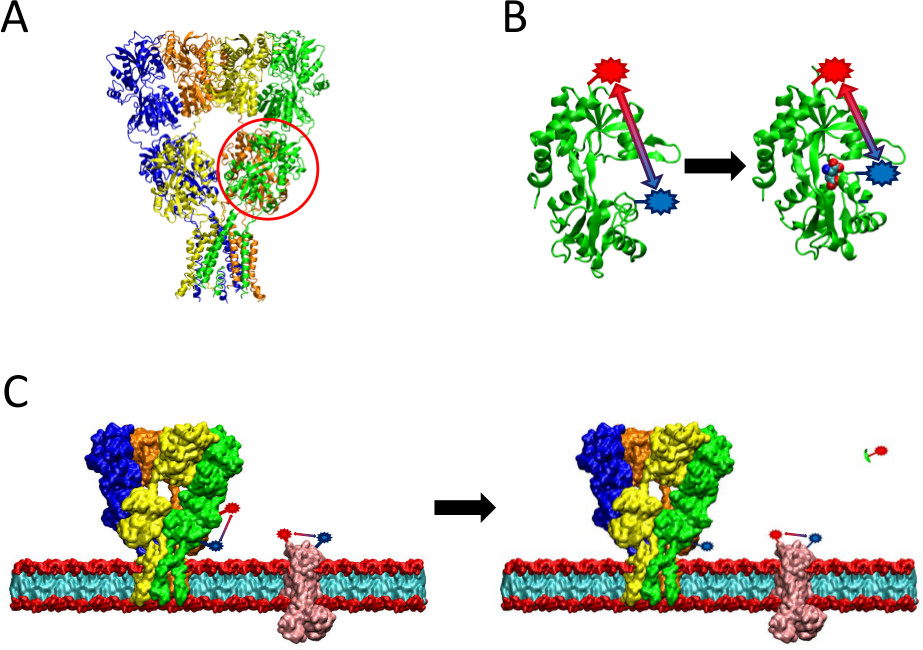
Figur 2. En oversikt over LRET metoden presentert. (A) AMPA-reseptoren er et membranprotein som gjennomgår konformasjonsendringer ved ligand-binding. Clamshell-formet ligand-binding domenet er sirklet her i rødt. (B) Den ligand-bindende domene av AMPA når den ikke er bundet til protein eksisterer i en åpen konformasjon (til venstre). Når bundet til ligand glutamat, lukkes protein rundt sin ligand (høyre). Ved å plassere fluoroforene på probative steder på LBD, kan innholdet i denne konformasjonsendringen sees som avstanden mellom fluoroforene endringer, som da vil påvirke fluorescens levetid. (C) Ved merking av hele celler, kan merking av begge for proteinet av interesse, så vel som bakgrunnsmembranproteiner forekommer (til venstre). Etter spalting protease, vil LRET signal fra proteinet av interesse forsvinner på grunn av frigjøring av et oppløselig fragment, slik at bakgrunnssignal intakt (høyre). Denne bakgrunnssignal kan så bli subtrahert fra det rå signal.
