The images that this technique produces can be analyzed in a variety of ways to collect information surrounding what happens after cells have phagocytosed C. neoformans. As seen in Figure 1, there are approximately 100 macrophages that are either infected or uninfected. Each one of these macrophages gives the viewer an opportunity to study host-pathogen interactions on a very microscopic level. As Figures 2 and 3 show, there are different outcomes that can occur between a macrophage and yeast cell, and even between macrophage-macrophage. Macrophages can be scored as undergoing non-lytic exocytosis, cell-cell transfer, or any other cellular process that may occur. The timing of such events during the infection can also be ascertained based on the parameters that are set when the movie is being recorded.
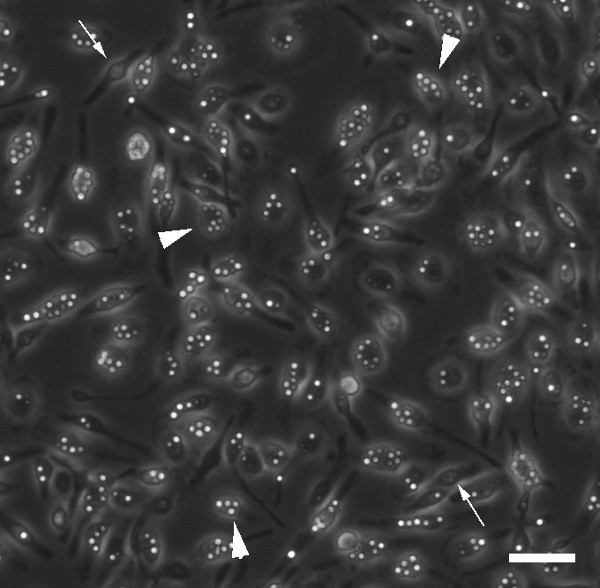
Figure 1: Field of Infected Macrophages. This figure shows an entire field of individually infected macrophages at the start of a 24 hr infection period. It should be noted that the field is free of extracellular Cn and more importantly, how clear it is which macrophages are infected (as indicated by a large arrowhead) and which contain no fungal cells (as indicated by a small arrow). Scale bar represents 50 μm.
Movie 1: Visualization of Macrophage-C. neoformans 24 hr Infection Period. The initial frame of the movie shows many macrophages throughout the field with some that are uninfected and some that have phagocytized varying amounts of cryptococcal cells. As the movie progresses, the cells are all motile and can be seen moving about the field. We are able to follow each of these macrophages to see what kind of cellular process they undergo.
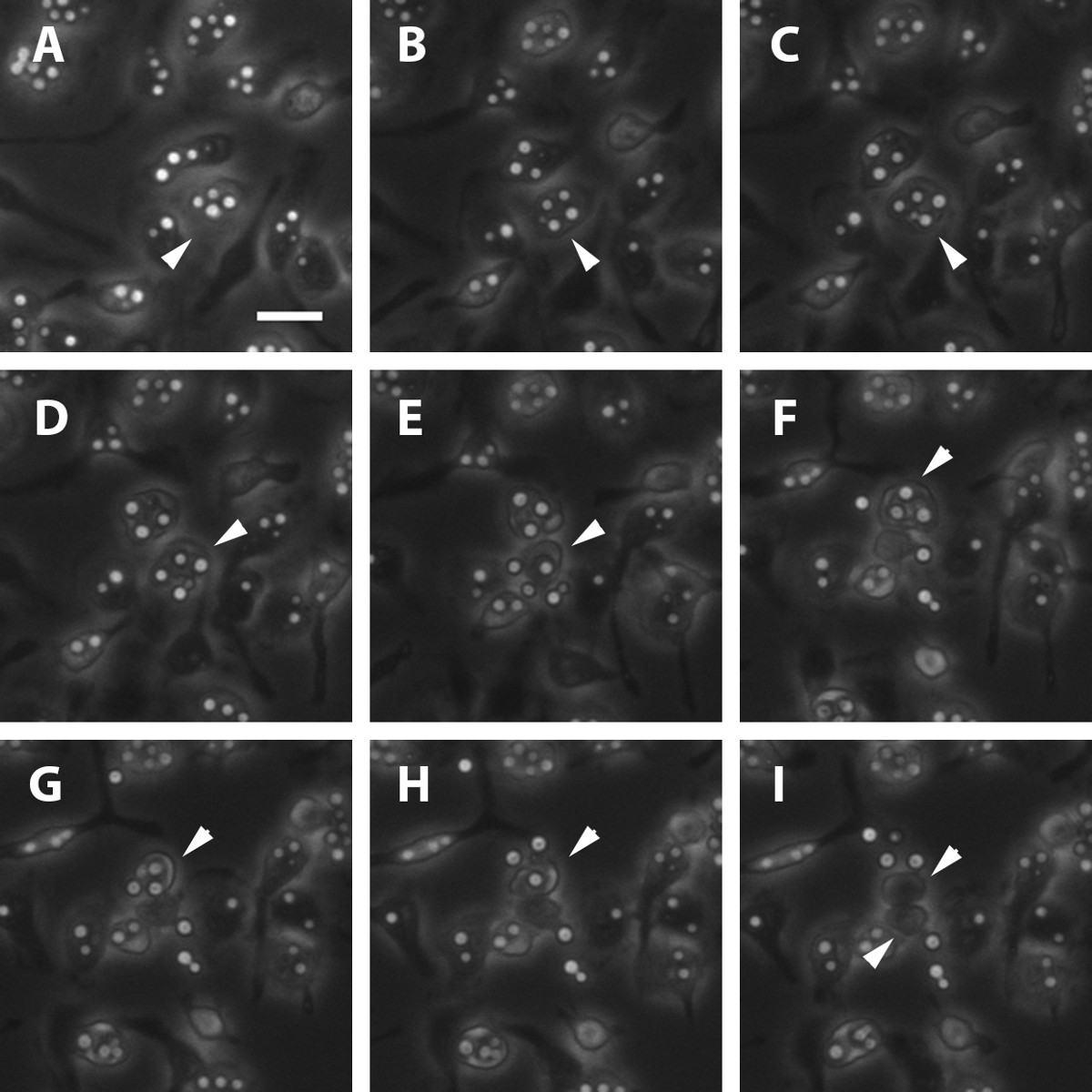
Figure 2: Primary Murine Macrophage Undergoing Non-lytic Exocytosis. An infected macrophage (Panel A, indicated by a white arrowhead) is seen undergoing the beginning stages of non-lytic exocytosis. The phagosome starts to enlarge (panel B-C) and the fungal load starts to shift. Several Cn cells are then released into the extracellular space as can be seen in panels D-E. A second infected macrophage in Panel F, indicated by a white arrowhead, also undergoes a non-lytic exocytosis event (Panel G-H). At the end of the process, both macrophages are left empty (Panel I) as shown by the white arrowheads. Scale bar represents 25 μm.
Movie 2: Visualizing Primary Murine Macrophage Undergoing Non-lytic Exocytosis. In this movie, it is clear that the cryptococcal cells are within the confines of an infected macrophage. The macrophage becomes agitated and a yeast cell can be seen budding within the phagosome. Cryptococcal cells are then quickly expunged into the surrounding environment. A second infected macrophage undergoes non-lytic exocytosis and more yeast cells are released into the extracellular space. The host cells remain viable after cryptococcal exocytosis as indicated by their ability to continue moving.
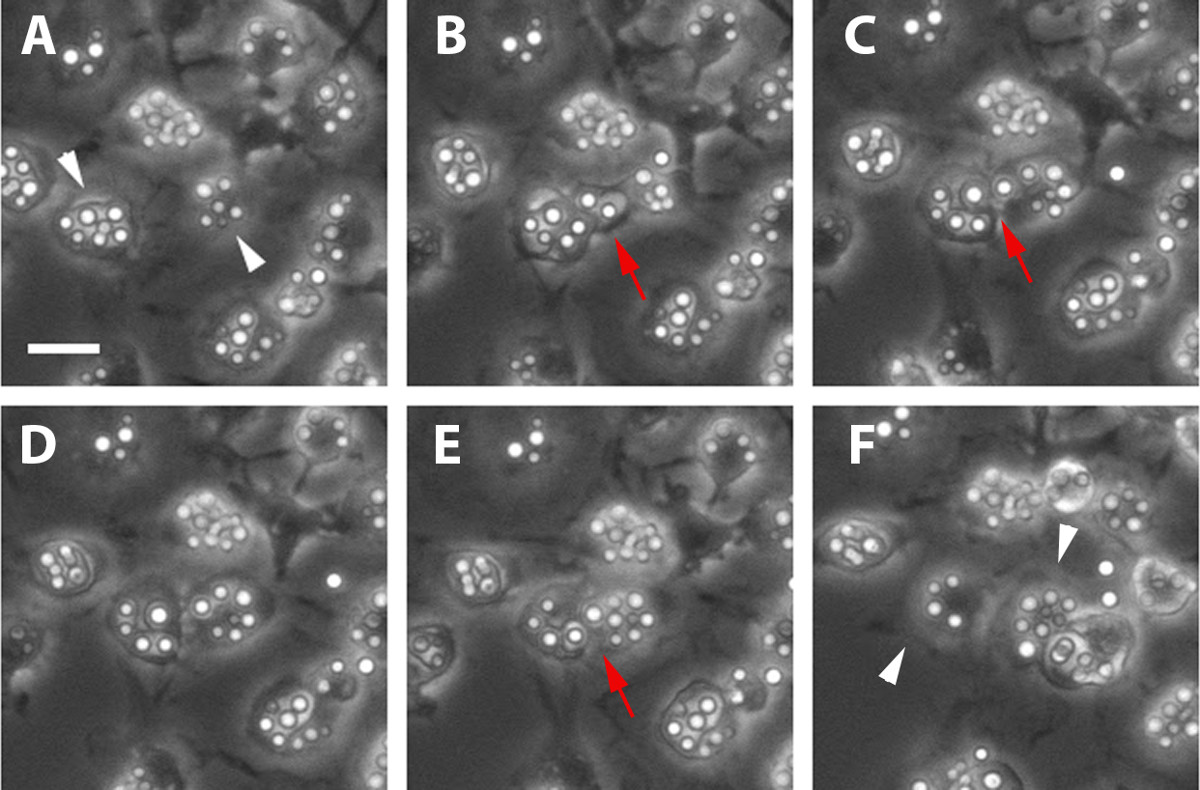
Figure 3: Primary Murine Macrophage Undergoing Cell-Cell Transfer. Another process that can be captured using this protocol is cell-cell transfer which occurs when cryptococcal cells are transferred from one macrophage to a neighboring macrophage. Two infected macrophages are in close proximity to each other indicated by white arrowheads (Panel A) and move even closer to initiate cell-cell transfer (Panel B). A cell-cell bridge indicated by the red arrowheads is formed between the two macrophages such that the cryptococcal cells are not exposed to the surrounding environment during the transfer (Panel C, E). After the transfer is complete, the cells break away (Panel D, F). Scale bar represents 25 μm.
Movie 3: Visualizing Cell-Cell Transfer Between Infected Macrophages. On occasion, macrophages will interact in such a way that facilitates transfer of cryptococcal cell between host cells. The yeast cells are not exposed to the extracellular environment and the host cells remain viable. In this movie, two infected macrophages create cell-cell bridges on two separate occasions to shuttle cryptococcal cells from one macrophage to another.
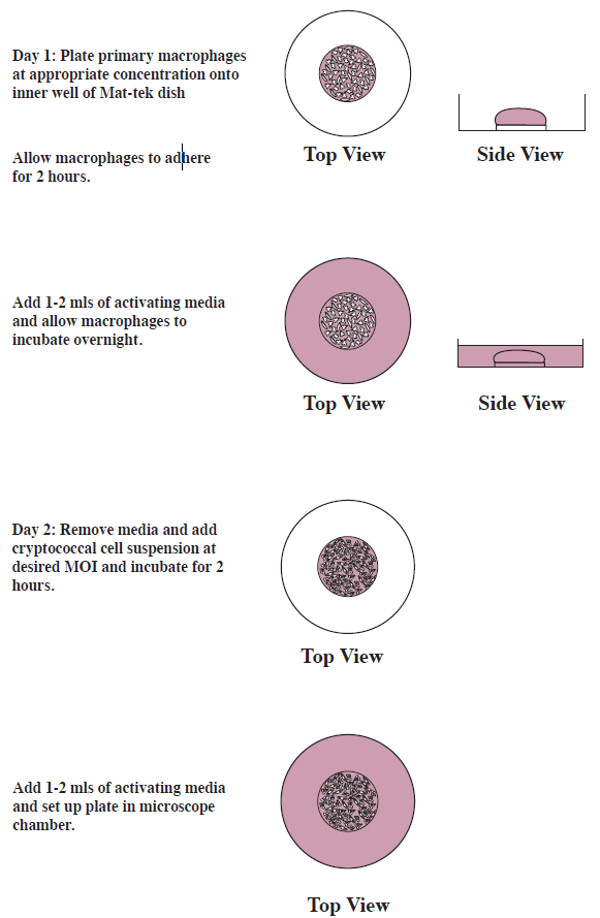
Figure 4: Schematic showing preparation of cells.
