A. gambiae:
Chitosan/dsRNA nanoparticles are formed due to the electrostatic interaction between amino groups of chitosan and the phosphate groups of dsRNA. The efficiency of dsRNA incorporation into nanoparticles is usually above 90% as measured by depletion of dsRNA from the solution. Atomic force microscopy images show that chitosan-dsRNA particle size averages 140 nm in diameter, ranging from 100-200 nm (Figure 1).

Figure 1. Nanoparticle formation by chitosan and interfering RNA. Atomic force microscopy image of chitosan/dsRNA nanoparticles (A) or chitosan solution without the addition of dsRNA (B). The size of the images was 1.0 μm × 1.0 μm. Scale bar = 250 nm. Please click here to view a larger version of this figure.
Feeding of these particles which are combined with standard larval food and incorporated into agarose supports normal larval development. Importantly, specific knockdown of ectoderm-derived AgCHS1 transcripts as well as midgut-specific AgCHS2 (Figure 2) is possible19, demonstrating that dsRNA taken up through ingestion of nanoparticles initiates RNAi beyond the midgut. Knockdown efficiencies ≈40-60% were observed (e.g., Figure 2) and vary between transcripts.
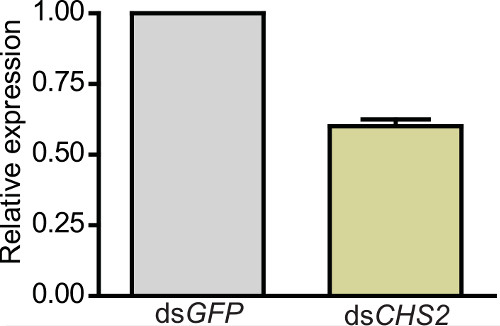
Figure 2.Transcription levels of chitin synthase 2 (AgCHS2) after dsRNA ingestion in A. gambiae larvae. Relative mRNA levels of AgCHS2 in larvae fed on nanoparticles with dsAgCHS2 or control dsGFP. Data are presented as mean ± SD from three independent biological replicates. Different letters on the bars indicate significant differences based on paired t tests (P = 0.003).
Knockdown of AgCHS1 significantly reduced the total chitin content of fourth instar larvae and increased the susceptibility to the chitin synthesis inhibitor insecticide, diflubenzuron19, Knockdown of AgCHS2 significantly increased the effect of calcofluor white and dithiothreitol which disrupted the peritrophic matrix (PM) in fourth instar larvae (Figure 3) resulting in increased mortality17.
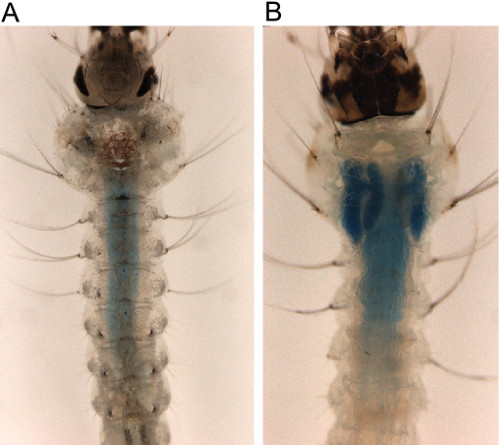
Figure 3. Impact of chitosan/dsAgCHS2 ingested nanoparticles on A. gambiae peritrophic matrix (PM) permeability. Calcofluor white or DTT treatment disrupts the PM of A. gambiae larvae which is further exuberated by AgCHS2 knockdown through ingestion of the chitosan/dsRNA nanoparticles19. (A) Mosquito with intact PM. (B) Mosquito with disrupted PM, dextran blue leaking into the gastric ceacae.
A. aegypti:
Chitosan/siRNA nanoparticles were used to target the axon guidance gene semaphorin-1a (sema1a) during A. aegypti larval development20. Knockdown of sema1a was confirmed through in situ hybridization, which detected reduced levels of sema1a in ≈60% of the larval brains assessed and failed to detect sema1a expression in ≈40% of knockdown animal brains (Figure4C vs. B; yellow arrowhead indicates the antennal lobe). qRT-PCR assays performed on pooled whole animals revealed that this technique resulted in 32 ± 10% reduction in sema1a transcripts as compared to control-nanoparticle fed animals (Figure 4A; P<0.01 based on paired t test, N=5, where N is the number of biological replicates). Knockdown in the brain persisted through at least 24 h of pupal development, as evidenced by loss of anti-Sema1a antibody expression (Figure 4E1 vs. D1), which was used to identify animals with significant knockdown during larval and pupal olfactory phenotype analyses. Larval and pupal antennal lobe defects (quantified in Table 1) were detected in sema1a knockdown larvae and pupae (Figure 4E2, E3 vs. D2, D3). Loss of Sema1a expression resulted in glomeruli that are deformed (Figure 4E2 vs. D2, visualized with mAbnc82; overlays of both signals are shown in Figure4D3, E3). Comparable phenotypes were generated with two separate sema1a knockdown siRNAs, which helped to ensure that the defects observed were not the result of off-site targeting by either siRNA. The highest levels of knockdown were generated when the siRNAs were combined, a strategy that can be used to increase knockdown efficiency (see Mysore et al.20 for more details).
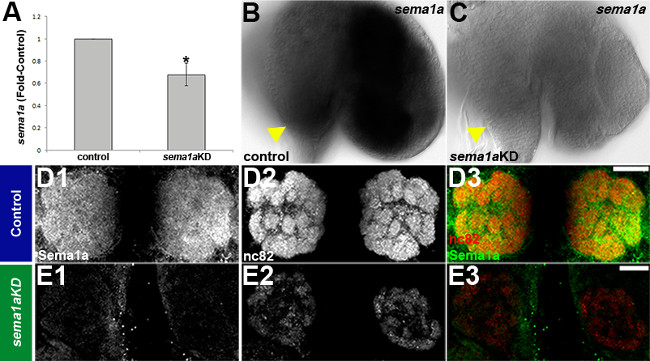
Figure 4. Chitosan/siRNA induced knockdown of sema1a in the developing olfactory system of A. aegypti. qRT-PCR (A) and in situ hybridization (B, C) assays confirmed knockdown of sema1a in larvae fed a mixture of sema1a targeting siRNA890 and siRNA1198 chitosan/interfering RNA nanoparticles vs. control nanoparticles. Loss of Sema1a expression resulted in glomeruli that are deformed (E1-3 vs. D1-3). See text for details. Dorsal is up in all panels. Scale bar = 25 μm. This figure originally appeared in Mysore et al20. Please click here to view a larger version of this figure.
| Larvae | Pupae | |||||
| siRNA | n | Wildtype | AL defects | n | Wildtype | AL defects |
| Control | 69 | 69 (100%) | 0 (0%) | 68 | 68 (100%) | 0 (0%) |
| sema1a KD | 77 | 42 (54.5%) | 35 (44%)* | 75 | 51 (68%) | 24 (32%)* |
Table 1. Quantification of antennal lobe defects following sema1a knockdown. A compiled summary of results obtained for control siRNA vs. sema1a knockdown (KD) siRNA890+siRNA1198 chitosan nanoparticle-fed animals from a total of eight replicate experiments performed in both larvae (left) and pupae (right) is shown. The total number of individuals assessed (n) and numbers/percentages of animals displaying wild type morphology vs. antennal lobe (AL) defects (olfactory receptor neuron targeting defects, defective neuropil and glomeruli formation) are indicated. Knockdown was verified immunohistochemically with anti-Sema1a antibody staining in a subset of animals (15 larvae and 10 pupae) that displayed the most severe defects (denoted with *). All knockdown animals evaluated in this manner were found to have reduced levels of Sema1a, while Sema1a levels in control-fed animals were not noticeably altered. This table originally appeared in Mysore et al20.
In a second investigation21, chitosan/siRNA nanoparticles were used to target the transcription factor Single-minded (Sim) during development of the olfactory system. qRT-PCR assays with pooled brains dissected from whole animals indicated that sim knockdown brains had on average a 47% reduction in sim transcripts in comparison to control nanoparticle-fed animals (P = 0.005 based on paired t test). Comparable assays with antennae revealed on average a 77% reduction in sim levels with respect to control-fed animals (P = 0.002 based on paired t test). Despite some variability in the levels of knockdown between tissues and animals, sim transcripts were not detected through in situ hybridization in ≈50% of the knockdown animal brains/antennae following treatment with either of two different knockdown siRNAs (Figure 5B2, B3, C2, C3 vs. B1, C1, Table 2). In yeast behavioral assays during which individuals that were attracted to yeast were awarded a score of 1, while animals that were not attracted were given a score of 0, average scores for sim430or sim718 knockdown animals were significantly lower than that of control-fed animals in two replicate experiments (Figure 5A; P<0.001 based on paired t test). Defective odor-tracking behavior (Figure 5A) correlated with reduced levels of sim in the antennae (Figure 5B2, B3 vs. B1) and brains (Figure 5C2, C3 vs. C1, yellow dots mark the perimeter of the antennal lobe; see Table 2 for quantification of results). Multiple defects were identified in the antennae and antennal lobes of sim knockdown animals (see Mysore et al. 21 for details). For example, sim knockdown larvae and pupae lacked expression of orco, the obligate co-receptor for all odorant receptors in olfactory receptor neurons (Figure 6). Reduced orco transcript levels were detected in individuals fed with sim430(Figure 6A3, B3) or sim718(Figure 6A4, B4) knockdown (KD) chitosan/siRNA nanoparticles (compare to wild type animals in Figure 6A1, B1 and control chitosan/siRNA nanoparticle-fed animals in Figure 6A2, B2). The loss of orco expression phenotype observed in L4 larvae (Figure 6A1-A4) persisted through at least 24h after pupal formation (Figure 6B1-B4).
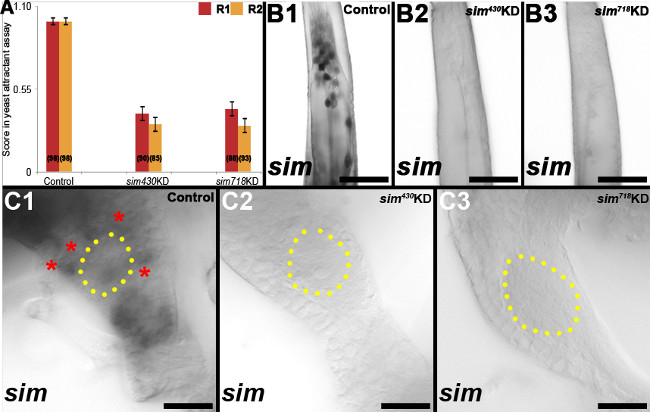
Figure 5. sim knockdown larvae show defective odor-attractant behavior. In situ hybridization revealed reduced levels of sim RNA in the antennae (B2, B3) and brains (C2, C3) of sim430and sim718 knockdown animals (compare to control-fed in B1, C1), which failed to respond to yeast odorant attractant (A). See text for details. The proximal ends of antennae are oriented upwards in B1-B3. Dorsal is oriented upward in C1-C3. Scale bar = 25 μm. This figure originally appeared in Mysore et al21. Please click here to view a larger version of this figure.
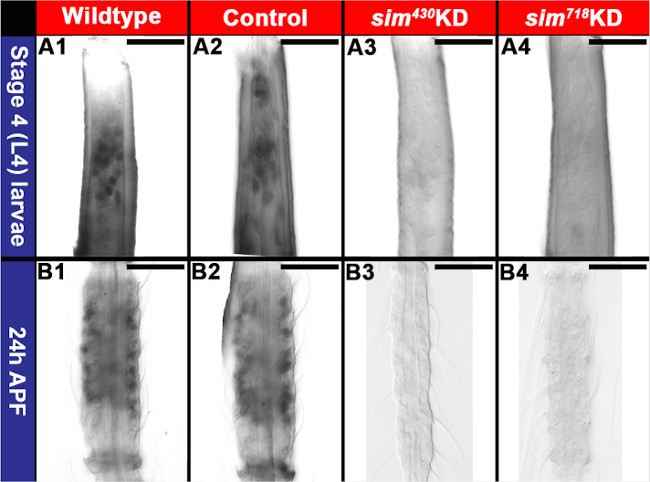
Figure 6. Loss of orco expression in the developing A. aegypti antenna following knockdown of sim. Reduced levels of orco transcript were detected in individuals fed with sim430 (A3, B3) or sim718 (A4, B4) knockdown (KD) chitosan/siRNA nanoparticles (compare to wild type animals in A1, B1 and control chitosan/siRNA nanoparticle-fed animals in A2, B2). See text for details. Scale bar = 25 μm. This figure was compiled from images published in Mysore et al21. Please click here to view a larger version of this figure.
| Attracted | Not Attracted | ||||||||
| siRNA | n | # Animals | Normal | Moderate | Null | # Animals | Normal | Moderate | Null |
| Control | 195 | 196 (100%) | 196 (100%) | 0 | 0 | 0 | 0 | 0 | 0 |
| sim430KD | 176 | 63 (35%) | 48 (76%) | 15 (24%) | 0 | 113 (64%) | 20 (18%) | 12 (11%) | 81 (71%) |
| sim718KD | 177 | 66 (37%) | 44 (66%) | 22 (34%) | 0 | 111 (63%) | 20 (18%) | 9 (8%) | 82 (74%) |
Table 2. Levels of sim correlate with larval performance in a yeast behavioral assay. A compiled summary of results obtained in four yeast odorant attractant replicate experiments is shown. The total number of animals (n) indicates the number of individual larvae that were tested in these behavioral assays. The number of individual larvae (# Animals) that were attracted (left; animals that touched the yeast pellet and were awarded a score of 1) or not attracted (right; animals that did not touch the yeast pellet and received a score of 0) under each condition (Control, sim430 KD, or sim718KD chitosan/siRNA nanoparticle-fed) are shown, and the percentages of total animals are included following the raw numbers. Knockdown of sim was assessed through in situ hybridization in the brains and antennae of animals attracted (left) or not attracted (right) to the yeast. The raw number/percentage of individuals with Normal (comparable to wildtype sim transcript levels), Null (no observable sim transcript), or Moderate (reduced but not wild type) sim levels are indicated. Loss of sim correlated well with a lack of attraction to the yeast. This table originally appeared in Mysore et al21.
