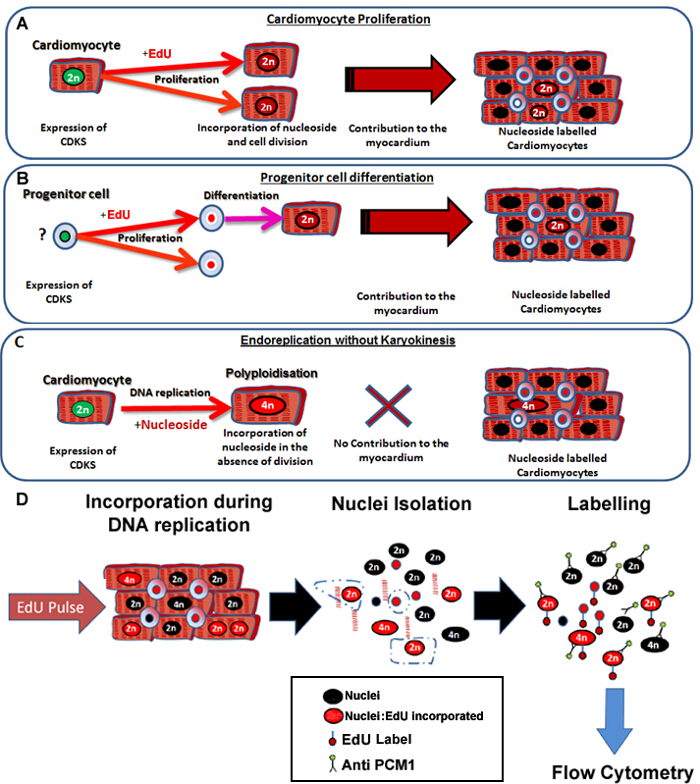
Animal arbejde blev godkendt og godkendt af Newcastle University Ethics Review Board. Alle dyreforsøg blev udført i overensstemmelse med retningslinjerne fra direktiv 2010/63 / EU af Europa-Parlamentet om beskyttelse af dyr, der anvendes til videnskabelige formål. 1. edu Administration Opløs edu i steril saltvandsopløsning (0,9% w / v) i en endelig koncentration på 12,5 mg / ml. Opvarm opløsningen til 40 ° C og vortex for at opløses fuldstændigt. Opbevar nok edu / saltvand ved 4 ° C for at injicere alle mus i undersøgelsen giver mulighed for 6 daglige injektioner. Bemærk: Typisk over 6 mus er nødvendige for hver forsøgsgruppe. Dog bør udføres styrkeberegninger for uafhængige undersøgelser. Afvej individuelle mus. Beregn den nødvendige volumen til at administrere 100 ug / g edu / saltopløsning til hver mus (8 pi lager edu opløsning pr g). Tegn en passende volume af edu / saltvand i insulinsprøjte med en 25 G nål. Udfør intraperitoneal (ip) injektioner mens håndtering mus ved hjælp af godkendte hjemmekontor teknikker. Placer en mus på buret låget. Træk forsigtigt tilbage på haleroden og fast forstå musen ved harmoniske. Udsætte maven til injektion ved at holde musen med pegefingeren og tommelfingeren nær bunden af hovedet og deponering dyrets hoved tilbage mod gulvet. Tør maven med 70% alkohol opløsning. Find dyrets midterlinjen og nederste højre kvadrant. Sprøjt edu / Saltvandsopløsning i dyret ind i den nederste højre kvadrant. Sprøjt mus med Edu / saltopløsning og registrere tidspunktet på dagen. Gentag trin 1,2 til 1,4 på samme tidspunkt på dagen, i 6 på hinanden følgende dage. Bemærk: Som der kræves en edu negativ kontrol for at indstille gating til flowcytometrisk analyse, injicere en ekstra alder, køn og eksperimentelt matchered mus med et tilsvarende volumen af saltvand uden edu. Dette vil give den nødvendige kontrol (ingen edu kontrol). Denne kontrol bør medtages for hver undersøgelse for at give mulighed for variationer i partier af reagenser og cytometer oprettet. 2. Høst Hjerter På 24 timer efter det 6. edu injektion, ofrer mus ved hjælp af cervikal dislokation (eller en alternativ Schedule 1 Home Office godkendt metode). Placer ofret dyr i rygleje og gøre huden indsnit fra midten maven til membranen med kirurgiske sakse. Skær mellemgulvet, og holde brystbenet væk fra kropskaviteten, skåret bilateralt at blotlægge hjertet. Løft hjerte lidt, og skære de store blodkar i udstrømningen tarmkanalen at dissekere hjertet ud af brysthulen. Anbring straks hvert hjerte i til en individuel 15 ml centrifugerør indeholdende 10 ml PBS nedkøles til 4 ° C. Overfør hjerter til SEParate 10 cm petriskåle under anvendelse af en skalpel skæres hvert hjerte i to i et sagittalt orientering og vask hjertet ved forsigtigt at klemme med en pincet. Derefter udskiftes PBS for at fjerne så meget blod som muligt. Gentag dette trin to gange. Placer begge dele af hver hjerte i det samme individ 1,5 ml mikrocentrifugerør. Fortsæt til trin 3 eller alternativt gem prøver -80 ° C som beskrevet i trin 2.9 og 2.10. Anvendelse af en 25 G nål, lave et lille hul i mikrocentrifugerør låg for at forhindre låget i at åbne som følge af ekspansionen af gasser når hjertet er behandlet. Mærk hver mikrocentrifugerør og sted i en beholder med flydende nitrogen til at fryse. 3. Nuclei Dissociation og cardiomyocyte Kerner Mærkning Bemærk: Denne kerner dissociation og cardiomyocyte kerner mærkning er tilpasset sig fra Bergmann et al 11 Udfør denne procedure på alle prøver injiceret.med Edu og saltvand injiceres (ingen edu kontrol) negativ kontrol. Se tabel 1 for opskriften af løsninger, der kræves for at udføre disse trin. For hvert hjerte, der skal analyseres, anbringes 36 ml 1% BSA / PBS-opløsning i en ultracentrifuge rør, inkuberes i 30 minutter, kasseres opløsningen og derefter lad det lufttørre. Bemærk: Dette rør anvendes i trin 3.6. Placer individuelle frosne hjerter på en 10 mm skål på is og hakkekød ved hjælp af en skalpel tillader hjerter at afrime under hakningen processen. Placer hakket hjerter i 15 ml Cell Lysis Buffer i et 50 ml centrifugerør og homogeniseres med en probe homogenisator i 15 sekunder ved 25.000 rpm ved stuetemperatur. Tilføj yderligere 15 ml Cell Lysis Buffer til centrifugerør og overførsel til en 40 ml Dounce med en stor clearance støder. Udfør 10 slag med pistil og filtreres gennem en 100 um derefter 40 um celle si i et 50 ml centrifugerør. Centrifuger i 10 minutter ent 700 xg ved 4 ° C og supernatanten fjernes. Resuspender nukleare pellet i 25 ml saccharosegradient Solution og lag cellekernerne indeholdende opløsning på toppen af 10 ml frisk saccharosegradient opløsning i ultracentrifugerør fremstillet i 3.1. Centrifuger ved 13.000 xg i 1 time ved 4 ° C ved anvendelse af en swing out rør rotoren. Efter centrifugering kasseres supernatanten og resuspender kerner pellet i 1.300 pi kerner opbevaring buffer i et 1,5 ml mikrocentrifugerør og etiket PCM1. Fjern 300 pi af suspensionen til en ny mikrocentrifugerør og tilføje til 700 pi frisk kerner opbevaringsbuffer. Mærk dette rør som ISO-kontrol. Tilføj anti-PCM1 ved en endelig koncentration på 8 pg / ml til mikrocentrifuge rør mærket PCM1 og tilføj Rabbit IgG isotype kontrol antistof i en koncentration på 8 pg / ml til mikrocentrifugerør mærket ISO kontrol. Inkuber natten over ved 4 ° C. Efter incubation, centrifugeres ved 700 xg i 10 min, supernatanten, erstatte med 1 ml frisk kerner opbevaring buffer, resuspender og centrifuger igen i 10 min ved 700 x g. Supernatanten kasseres og erstattes med 1 ml frisk kerner opbevaringsbuffer. Tilsæt 1 pi F (ab ') 2-fragment af gede-anti-kanin IgG (H + L) antistof (FITC eller tilsvarende grønt-fluorescerende farvestof) for at opnå en slutkoncentration på 2 pg / ml. Bemærk: Brugen af et F (ab ') 2-fragment-antistof reducerer potentialet for ikke-specifik mærkning af immunceller, herunder makrofager og B-celler. Wrap rør i aluminiumfolie for at beskytte mod lys og inkuberes i 1 time ved 4 ° C. 4. Påvisning af Edu Indbygning Fortynd 10X koncentreret edu reaktionsbuffer 1:10 med deioniseret vand. Centrifuger PCM-1 og ISO kontrol mærkede kerner suspensioner ved 700 xg i 10 min, supernatanten og resuspender kerner pellet i 1 ml1% BSA / PBS-opløsning. Vask kerner pellet to gange. Efter den sidste vask, kasseres supernatanten og erstatte med 100 ul edu fiksativ. Wrap rør i aluminiumfolie for at beskytte mod lys og inkuber ved stuetemperatur i 15 minutter. Vask prøverne to gange med 1 ml 1% BSA / PBS-opløsning, centrifugeres ved 700 xg og inkuberes med 1x saponin-baserede permeabilisering opløsning ved stuetemperatur i 15 minutter. Forbered 1x edu mærkning cocktail (tabel 2). Et minimum af 2 reaktioner vil være forpligtet til også at omfatte ikke edu kontrol. Der tilsættes 500 pi 1x edu mærkning cocktail direkte til hver prøve, der allerede indeholder kerner i 100 pi 1x saponin-baserede permeabilisering opløsning og inkuber ved stuetemperatur beskyttet mod lys i 30 minutter. Centrifuger ved 700 xg og resuspender i 1 ml 1% BSA / PBS og gentage dette trin to gange mere. Centrifuger ved 700 xg, kasseres supernatanten og erstatte med 400 pi DNA farveopløsning (se tabel of Materials). Navn af reagenset 1. Cell Lysis Buffer 0,32 M saccharose 10 mM Tris-HCl (pH = 8) 5 mM CaCl2 5 mM magnesiumacetat 2,0 mM EDTA 0,5 mM EGTA 1 mM DTT 2.Sucrose Gradient Solution 2,1 M saccharose 10 mM Tris-HCl (pH = 8) 5 mM magnesiumacetat 1 mM DTT 3. Kerner storage buffer (NSB) 0,44 M saccharose 10 mM Tris-HCI (pH = 7,2) 70 mM KCI 10 mM MgCl2 1,5 mM spermin Tabel 1: kerner isolation buffer ingredienser Opskrift på alle buffere, der anvendes i protokol afsnit 3 (Nuclei dissociation og cardiomyocyte kerner mærkning).. 5. flowcytometrisk analyse Bemærk: Udfør Flow cytometri på isolerede kerner som tidligere 7,11,15 beskrevet. Opret først et plot, der beskriver Forward Scatter (FSC) og Side Scatter (SSC) at tillade diskrimination af kerner fra affald (figur 2A). Opret et plot til at skelne enkelte kerner fra aggregater (figur 2B) ved at plotte 4 ', 6-diamidino-2-phenylindol) (DAPI) område (3-405 / 450/50-A) versus højde (33-405 / 450 / 50-H). Sørg that 3-405 / 450/50 Et signal opsamles på den lineære skala for at muliggøre korrekt bestemmelse af DNA-indhold. Opret et plot, der beskriver 488/535/30-A (PCM-1) vs SSC og viser kun begivenheder fra singlet gate skabt i 5.2 for at vurdere PCM-1-ekspression i singlet befolkning. Kør Isotype kontrolprøven for at etablere en PCM-1 positive gate (figur 2C). Køre en lille mængde PCM1 mærkede prøve at kontrollere PCM1 udtrykkende befolkning og gating position. Opret et plot, der beskriver SSC-A vs 405/450/50-A (at opdage DAPI). I dette plot, display kun singlet PCM1 udtrykker kerner, ved hjælp af portene oprettet i 5.3. Brug denne plot til at oprette en ekstra hierarkiske port, der kun indeholder 2 N kerner (figur 2D). Opret et plot, der beskriver 488/535 / 30-A vs 1-633 / 660/20-A for at tillade identifikation af Edu mærkning PCM1 udtrykke cardiomyocyte befolkning. Vis kun kerner, som er singlet, P CM1 + og 2N hjælp af ovenstående gating. Kør hjerte prøve fra nogen Edu kontrol for at indstille edu positive port (Figur 2E). Opret passende gate for Edu + celler. Optag og kvantificere singlet, PCM-1 udtrykker, 2N kerner, der har indarbejdet Edu. Beregn denne population som en procentdel af den samlede singlet, PCM1 + kerner. Dette vil give hastigheden af neo-cardiomyocyte kerner generation, i løbet af syv dage edu puls periode, som en procentdel af de samlede cardiomyocyte kerner. Bemærk: Som DAPI binding til DNA er støkiometrisk fluorescensintensiteten er proportional med mængden af DNA. Bestem ploidi baseret på intensiteten af 405/450/50 Et signal, som tidligere 7 beskrevet. At evaluere ploidi inden for den samlede cardiomyocyte befolkning bruger plottet etableret i 5.5 og en gating strategi baseret på DNA-koncentrationen 7. /files/ftp_upload/53979/53979fig2.jpg "/> Figur 2:. Gating strategi at kvantificere cardiomyocyte Edu inkorporering og ploidi (A) Kerner diskrimineres fra snavs baseret på forward scatter (FSC) og sidespredning (SSC). (B) En lineær port er skabt og den singlet kerner befolkning er identificeret baseret på DAPI mærkning og 405/450/50 højde vs område signal. (C) Fluorescerende gating tillader adskillelse af cardiomyocyte kerner (PCM-1-positive) og ikke-cardiomyocyte (PCM-1-negative) kerner fra hjertevævet. (D) Intensitet af DAPI signal anvendes til at bestemme DNA-koncentration og dermed kerner Ploidiniveauet for PCM1 + cardiomyocyte befolkning. I mus> 80% af cardiomyocyte kerner er diploide (2n). (E) Fluorescerende gating tillader adskillelse af 2N, cardiomyocyte kerner som har inkorporeret edu (PCM + / EDU + = 0,9%) fra 2 N, cardiomyocytter, som ikke har inkorporeret edu (PCM + uddan-). Alle trin er beskrevet i protokollen 5.0. Eksempel vist fra MDX -. / Å mus på C57 / BL10 baggrund Klik her for at se en større version af dette tal.