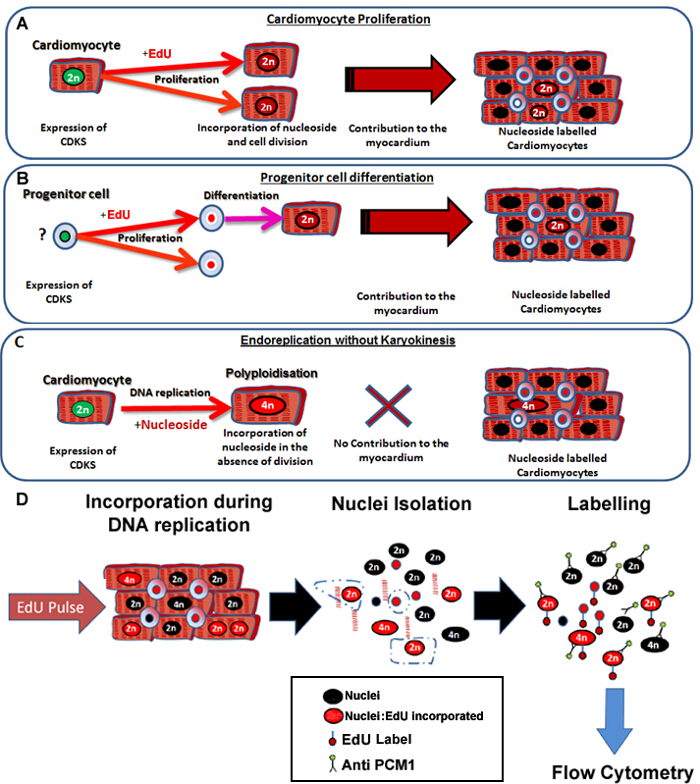
Animal arbeidet ble autorisert og godkjent av Newcastle University Etikk Review Board. Alle dyr prosedyrer ble utført i samsvar med retningslinjene fra direktiv 2010/63 / EU fra Europaparlamentet om beskyttelse av dyr som brukes til vitenskapelige formål. 1. Edu Administrasjon Oppløs EDU i steril saltoppløsning (0,9% vekt / volum) ved en sluttkonsentrasjon på 12,5 mg / ml. Oppvarm løsningen til 40 ° C og vortex for å fullstendig oppløse. Oppbevar nok Edu / Saline ved 4 ° C for å injisere alle musene i studien åpner for 6 daglige injeksjoner. Merk: Vanligvis over 6 mus er påkrevd for hver forsøksgruppe. Imidlertid bør beregninger utføres for uavhengige studier. Vei individuelle mus. Beregn nødvendig volum for å administrere 100 mikrogram / g Edu / saltløsning til hver mus (8 mL av lager Edu løsning per g). Tegn riktig volume av Edu / saltvann inn insulin sprøyte med en 25 G nål. Utfør intraperitoneal (ip) injeksjoner ved håndtering av mus ved bruk godkjente hjemmekontoret teknikker. Plasser en mus på buret lokket. Trekk forsiktig tilbake på haleroten og fast grep musen ved Scruff. Expose magen for injeksjon ved å holde musen med pekefingeren og tommelen nær bunnen av hodet og tipping dyrets hode bakover mot gulvet. Tørk magen med 70% alkohol. Finn dyrets midtlinjen og nedre høyre kvadrant. Injiser Edu / Saline løsning i dyret inn i den nedre høyre kvadrant. Injisere mus med Edu / saltvannsoppløsning og ta den tiden av døgnet. Gjenta trinn 1,2 til 1,4 på samme tid på dagen, 6 dager. Merk: Som en Edu negativ kontroll er nødvendig for å sette gating for flowcytometrisk analyse, injisere en ekstra alder, kjønn og eksperimentelt matcheed mus med et tilsvarende volum av saltvann uten Edu. Dette vil gi den nødvendige kontrollen (uten Edu kontroll). Denne kontrollen bør inkluderes for hver studie for å gi rom for variasjoner i grupper av reagenser og cytometer satt opp. 2. Høsting Hearts På 24 timer etter den 6. Edu injeksjon, ofre musene bruker halshugging (eller en alternativ Schedule en Home Office godkjent metode). Plasser ofret dyr i liggende stilling og gjør huden snitt fra midten av magen til membranen med kirurgiske saks. Skjær membranen, og holder sternum vekk fra kroppens hulrom, kuttet bilateralt å avsløre hjertet. Løft hjertet litt og kutte de store blodårene i utstrømningen kanalen å dissekere hjertet ut av brysthulen. Umiddelbart hvert hjerte i til en individuell 15 ml sentrifugerør inneholdende 10 ml PBS avkjølt til 4 ° C. Overfør hjerter til oljarate 10 cm petriskåler, ved hjelp av en skalpell kuttes hvert hjerte i to i en sagittal orientering og vaske hjertet ved forsiktig klemme med en pinsett. Deretter erstatte PBS for å fjerne så mye blod som mulig. Gjenta dette trinnet to ganger. Plassere begge delene av hvert hjerte i samme individ 1,5 ml mikrosentrifugerør. Fortsett til trinn 3 eller alternativt lagre prøvene -80 ° C som beskrevet i trinn 2,9 og 2,10. Ved å bruke en 25 G nål, lage et lite hull i mikrosentrifugerør lokk for å hindre at lokket fra å åpnes på grunn av utvidelsen av gasser når hjertet blir behandlet. Merk hver mikrosentrifugerør og plasser i en beholder med flytende nitrogen til å fryse. 3. Kjerner Dissosiasjon og kardiomyocytt Nuclei merking Merk: Denne kjerner dissosiasjon og cardiomyocyte kjerner merking er tilpasset fra at av Bergmann et al 11 Utfør denne prosedyren på alle prøvene injisert.med Edu og saltvann injisert (ingen Edu kontroll) negativ kontroll. Se tabell 1 for oppskriften til løsninger som kreves for å utføre disse trinnene. For hvert hjerte som skal analyseres, plasserer 36 ml av 1% BSA / PBS løsning i en ultrasentrifuge tube, inkuber i 30 min, skal den kastes, og deretter la det lufttørke. Merk: Dette rør blir brukt i trinn 3,6. Plasser individuelle frosne hjerter på en 10 mm rett på is og kjøttdeig ved hjelp av en skalpell slik at hjertene til tine under hakking prosessen. Plassere hakket hjerter i 15 ml Cell Lysis Buffer i et 50 ml sentrifugerør og homogenisere med en sonde homogenisator i 15 sekunder ved 25.000 rpm ved romtemperatur. Tilsett ytterligere 15 ml Cell Lysis Buffer til sentrifugerøret og overføring til en 40 ml Dounce med en stor klaring pistill. Utføre 10 slag med pistill og filtrer gjennom en 100 um deretter 40 mikrometer celle sil inn i et 50 ml sentrifugerør. -Sentrifuge i 10 min ent 700 xg ved 4 ° C og fjern supernatanten. Resuspender atom pelleten i 25 ml sukrosegradient løsning og lag cellekjerner inneholdende oppløsning på toppen av 10 ml frisk sukrose gradient løsning i ultrasentrifugerør fremstilt i 3.1. Sentrifuger ved 13000 x g i 1 time ved 4 ° C ved anvendelse av en sving ut rør rotor. Etter sentrifugeringen, kast supernatanten og resuspender pelleten kjerner i 1300 mL av kjerner lagringsbuffer i et 1,5 ml mikrosentrifugerør og etikett PCM1. Fjerne 300 pl av suspensjonen til et nytt mikrosentrifugerør og tilsett 700 ul av frisk kjerner lagringsbuffer. Merk dette røret som ISO-kontroll. Legg anti-PCM1 ved en endelig konsentrasjon på 8 ug / ml til mikrosentrifugerør merket PCM1 og legge til kanin IgG-isotype kontroll-antistoff i en konsentrasjon på 8 ug / ml til mikrosentrifugerør merket ISO-kontroll. Inkuber over natten ved 4 ° C. Etter incubasjon, sentrifuger ved 700 xg i 10 min, kast supernatanten, erstatte med 1 ml frisk kjerner lagring buffer, resuspender og sentrifuger igjen i 10 min ved 700 x g. Kast supernatanten og erstatte med 1 ml frisk kjerner lagring buffer. Tilsett 1 pl av F (ab ') 2 fragment av geit anti-kanin IgG (H + L) antistoff (FITC eller tilsvarende grønt-fluorescerende fargestoff) for å oppnå en sluttkonsentrasjon på 2 ug / ml. Merk: Bruken av et F (ab ') 2-fragment antistoff minsker potensialet for ikke-spesifikk merking av immunceller, inkludert makrofager og B-celler. Vikle rør i aluminiumfolie for å beskytte mot lys og inkuberes i 1 time ved 4 ° C. 4. Påvisning av Edu innlemmelse Fortynnet 10x konsentrert Edu reaksjonsbuffer 1:10 med avionisert vann. Sentrifuger PCM-1 og ISO kontroll merkede kjerner suspensjoner ved 700 xg i 10 minutter, kaste supernatanten og resuspender pelleten kjerner i 1 ml1% BSA / PBS-oppløsning. Vask kjerner pellet to ganger. Etter siste vask, kast supernatanten og erstatte med 100 ul av Edu fiksativ. Vikle rør inn i aluminiumsfolie for å beskytte mot lys og inkuber ved romtemperatur i 15 min. Vask prøvene to ganger med 1 ml 1% BSA / PBS-løsning, sentrifuger ved 700 xg og inkuber med 1x saponin-baserte permeabiliseringen løsning ved romtemperatur i 15 min. Forbered 1x Edu merking cocktail (tabell 2). Et minimum av 2 reaksjoner vil være nødvendig å inkludere noen Edu kontroll. Legge til 500 ul av 1 x edu merking cocktail direkte til hver prøve som allerede inneholder atomkjerner i 100 pl 1 x saponin-baserte permeabiliseringen løsning og inkuber ved romtemperatur beskyttet mot lys i 30 minutter. Sentrifuger ved 700 xg og resuspender i 1 ml av 1% BSA / PBS og gjenta dette trinnet to ganger til. Sentrifuger ved 700 xg, kast supernatanten og erstatte med 400 mL av DNA fargeløsning (se tabell for material). Navn på reagens 1. Cell Lysis Buffer 0,32 M sukrose 10 mM Tris-HCl (pH = 8) 5 mM CaCl2 5 mM magnesiumacetat 2,0 mM EDTA 0,5 mM EGTA 1 mM DTT 2.Sucrose Gradient Solution 2,1 M sukrose 10 mM Tris-HCl (pH = 8) 5 mM magnesiumacetat 1 mM DTT 3. Nuclei lagring buffer (NSB) 0,44 M sukrose 10 mM Tris-HCl (pH = 7,2) 70 mM KCl 10 mM MgCl2 1,5 mM spermin Tabell 1: kjerner isolasjon buffer ingredienser Resept for alle Buffere brukes i protokollen § 3 (Kjerner dissosiasjon og kardiomyocytt kjerner merking).. 5. flowcytometrisk analyse Merk: Utfør Flowcytometri på isolerte kjerner som beskrevet tidligere 7,11,15. Først oppretter en tomt som beskriver Forward Scatter (FSC) og Side Scatter (SSC) for å tillate diskriminering av kjerner fra rusk (figur 2A). Lag en plan for å diskriminere enkeltkjerner fra aggregater (figur 2B) ved å plotte 4 ', 6-diamidino-to-phenylindole) (DAPI) område (3-405 / 450/50-A) versus høyde (33-405 / 450 / 50-H). sikre that 3-405 / 450/50 A signal blir oppsamlet på den lineære skala for å gi den korrekte bestemmelse av DNA-innholdet. Lag en tomt som beskriver 488/535 / 30-A (PCM-1) vs SSC og vise bare hendelser fra sing gate opprettet i 5.2 for å vurdere PCM-1 uttrykk i singlet befolkningen. Kjør Isotype kontrollprøven for å etablere et PCM-en positiv port (figur 2C). Kjør en liten mengde PCM1 merket prøve å verifisere PCM1 uttrykke befolkningen og gating posisjon. Lag et plot som beskriver SSC-A vs 405/450/50-A (for å oppdage DAPI). I denne tomten, vise bare singlet PCM1 uttrykker kjerner, ved hjelp av portene som er opprettet i 5.3. Bruk denne tomten for å opprette en ekstra hierarkisk gate som bare inneholder 2 N kjerner (figur 2D). Lag en tomt som beskriver 488/535 / 30-A vs 1-633 / 660/20-A for å tillate identifisering av Edu merking av PCM1 uttrykke kardiomyocytt befolkningen. Vis bare atomkjerner som er singlet, P CM1 + og 2N ved hjelp av den ovennevnte gating. Kjør hjerte prøve fra noen Edu kontrollen for å stille Edu positive gate (figur 2E). Skape den riktige porten for Edu + celler. Ta opp og kvantifisere singlet, PCM-1 som uttrykker, 2N kjerner som har innarbeidet Edu. Beregn denne populasjonen som en prosent av total singlet, PCM1 + kjerner. Dette vil gi frekvensen av neo-kardiomyocytt kjerner generasjon, i løpet av 7 dagers EDU puls periode, i prosent av totale cardiomyocyte kjerner. Merk: Når DAPI-binding til DNA er støkiometrisk fluorescens intensitet er proporsjonal med mengden av DNA. Bestemme ploiditet basert på intensiteten av 405/450/50-A-signal, slik som beskrevet tidligere 7. For å evaluere ploiditeten innen totale cardiomyocyte befolkningen bruke tomten etablert i 5.5 og en gating strategi basert på DNA-konsentrasjon 7. /files/ftp_upload/53979/53979fig2.jpg "/> Figur 2:. Avblending strategi for å kvantifisere kardiomyocytt Edu innlemmelse og ploiditet (A) Nuclei blir diskriminert fra rusk basert på fremover scatter (FSC) og side scatter (SSC). (B) En lineær port er opprettet, og sing kjerner populasjonen er identifisert basert på DAPI merkingen og den 405/450/50 høyde vs arealsignal. (C) Fluorescerende gating tillater separasjon av cardiomyocyte kjerner (PCM-1-positive) og ikke-cardiomyocyte (PCM-1-negativ) kjerner av hjertevevet. (D) Intensitet av DAPI signal anvendes for å bestemme DNA-konsentrasjon og derved kjerner ploiditet av PCM1 + kardiomyocytt befolkning. I mus> 80% av cardiomyocyte kjerner er diploid (2n). (E) Fluorescent gating tillater separasjon av 2N, cardiomyocyte kjerner som har innarbeidet Edu (PCM + / EDU + = 0,9%) fra 2N, cardiomyocytes som ikke har innarbeidet Edu (PCM + undervis-). Alle trinnene er beskrevet i protokollen 5,0. Eksempel vist fra MDX -. / Y mus på C57 / BL10 bakgrunn Vennligst klikk her for å se en større versjon av dette tallet.