Adenosin (ADO) er en purinnukleosidanalog med en adenin molekyl festet til et ribose sukkermolekyl gruppen via en glykosidbinding. Når den er tilstede i det ekstracellulære miljøet, beskytter cellene mot overdreven skade ved virkningen av immunsystemet. Denne rollen har blitt fremhevet ved hjelp av ulike sykdomsmodeller, for eksempel kolitt 1, diabetes 2, astma 3, sepsis 4, og iskemisk skade fem. En av hovedfunksjonene ADO er inhibering av immunresponsen hos svulsten mikromiljøet, noe som bidrar til reduksjon av tumorImmunflukt 6. Av denne grunn er de mekanismene som er involvert i dannelse og ADO signalering er av betydelig terapeutisk interesse 7.
ADO nivåer i vev mikro er forholdsvis lav under normale fysiologiske betingelser og i hvert fall under sensitivitetsgrensen på immunceller. Men under hypoksi, ischemi, inflammasjon, infeksjon, metabolskstress og svulst forvandling de raskt øke åtte. De forhøyede ekstracellulære ADO nivåer i respons til vev-perturbing signaler har en dobbel funksjon: å rapportere vev skade i en autokrint og parakrint måte og å generere vev responser som kan generelt sett på som cytoprotective.
Ekstracellulær ADO kan dannes gjennom en rekke mekanismer, som inkluderer frigivelse av intracellulære kamre mediert av nukleosid-transportører 9 eller akkumulering på grunn av nedsatt nedbrytning drives av adenosindeaminase. Den viktigste vei som fører til økte ekstracellulære nivåer ADO omfatter virkningen av en kaskade av ectonucleotidases, som er membran-assosiert ectoenzymes genererer ADO ved phosphohydrolysis av nukleotider som frigjøres fra døde eller døende celler. Denne veien går gjennom den sekvensielle handling av CD39 (ectonucleoside trifosfat diphosphohydrolase-1) som konverterer ekstracellulære adenosin 5'-trifosfat (ATP) eller adenosin-5'-difosfat (ADP) til adenosin-5'-monofosfat (AMP) og CD73 (5'-nukleotidase), som konverterer AMP til ADO 10.
Ekstracellulær ADO utløser sine fysiologiske responser ved å binde seg til fire transmembrane ADO-reseptorer, nemlig A1, A2A, A2B og A3. Hver reseptor har forskjellige affinitet for ADO og spesifikk fordeling vev. Alle reseptorene har syv transmembrandomener og er i G-proteinkoblet til intracellulære GTP-bindende proteiner (G-proteiner), som kan indusere (Gs-proteinet) eller inhiberer (Gi protein) adenylatsyklase-aktivitet og, senere, produksjonen av intracellulær cAMP. Derfor endringer i cytoplasma cAMP-nivåer innvirkning på intracellulært protein-kinase-aktivitet under fysiologiske responser 11. Under fysiologiske betingelser ekstracellulære ADO er under 1 pm, som kan aktivere ukritisk A1, A2A og A3-reseptorer. Imidlertid aktivering av A2B subtype krever betydelig høyerekonsentrasjoner av nukleosidet, for eksempel de som er oppnådd under patofysiologiske betingelser. Alternativt kan ekstracellulære ADO bli degradert til inosin (INO) av adenosindeaminase (ADA) og CD26, en ADA kompleksprotein lokalisere ADA på celleoverflaten. En annen mulighet er at ADO er internalisert av cellen gjennom nukleosidlikevektstransportør transportører (ØNH) og fosforylert til AMP av ADO kinase protein 12,13.
Formålet med denne protokollen er å beskrive en analytisk metode for revers fase høy-ytelse væskekromatografi (RP-HPLC) for å kvantifisere i en enkelt kjøring substratet AMP og produktene ADO og INO, som genereres av humane lymfocytter. Vår erfaring var i utgangspunktet oppnådd ved bruk av celler fra kronisk lymfatisk leukemi (KLL) pasienter, som er preget av utvidelsen av en moden befolkning på CD19 + / CD5 + B-lymfocytter konstitutivt uttrykker CD39 14,15. Vi viste ca 30%av KLL pasienter uttrykker CD73 ectoenzyme og at dette fenotype korrelerer med dårlig prognose 16. Denne undergruppe av leukemiceller co-uttrykker CD39 og CD73 kan aktivt produsere ekstracellulære ADO fra ADP og / eller AMP. Preinkubasjon av CD73 + CLL celler med α, β-metylen-ADP (APCP), en kjent inhibitor av CD73 enzymatisk aktivitet, fullstendig blokkerer ekstracellulære ADO syntese bekreftet at CD73 representerer en flaskehals og enzym fra det kaskade 16.
KLL celler også uttrykker ADA og ADA komplekse protein CD26, som er ansvarlig for konvertering av ADO i INO. Ved å bruke spesifikke ADA-inhibitorer, slik som erytro-9- (2-hydroksy-3-nonyl) I wiadenine (EHNA) -hydroklorid og deoksykoformycin (DCF), er det mulig å blokkere ekstracellulære ADO degraderingen INO. Videre forbehandling med en ADA-inhibitor i kombinasjon med dipyridamol, som blokkerer nukleosid transportører, forbedrer ADO akkumulering i cellensupernatanter.
Vi har deretter utvidet denne protokollen til celler som stammer fra andre linjer, inkludert T-lymfocytter og myeloide celler, bekrefter CD73-avhengig ADO produksjon. Disse funnene tyder på at denne HPLC-protokollen er meget allsidig, og at den kan brukes til forskjellige celle linjer og til forskjellige dyrkningsbetingelser (figur 1).
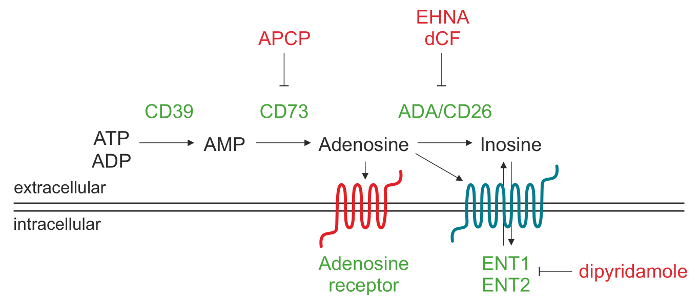
Figur 1. Skjematisk fremstilling av den enzymatiske maskineri ansvarlig for ekstracellulær ADO produksjon. Adenosin-5'-trifosfat (ATP) og / eller adenosin-5'-difosfat (ADP) kan brytes ned av CD39 til adenosin 5'-monofosfat (AMP), hvilken i sin tur omdannes av CD73 til nukleosidet adenosin (ADO). Når ADO er produsert i det ekstracellulære rom, kan det inn igjen i cellen gjennom nukleosid transportører (ENT), omdannes til inosin (INO) ellerbinde seg til forskjellige typer P1 ADO reseptorer. Klikk her for å se en større versjon av dette tallet.
