מסלולי העברת אותות לשחק תפקיד משמעותי כמעט בכל תהליך הסלולר המאפשרים לתא להגיב לאותות סביבתיים במהירות. 1 מסלולים אלה הם בדרך כלל מופעלים על ידי הקשירה של מולקולת איתות קולטן תאי, שתוצאתה הפעלה של אנזימים תאיים. הגברת התפשטות של האיתותים בתוך התא מתווכת על ידי הפונקציה של איתות חלבונים יוצרים רשת של אינטראקציות בין חלבונים שבו אנזימים מופעלים הפיך עם סגוליות גבוהות. בגלל חוסר ויסות של רשתות אלה לעיתים קרובות מוביל להתפתחות סרטן, חלה התעניינות רבה בהקמת 'טיפול הולכת אותות של סרטן', 2 לפיה תרופות נועדו לשבש מסלולי איתות ממאיר. הצענו לאחרונה גישה חלופית לאותת טיפול תמרה המסתמך על היכולת של תרופות כדי ליצור מסלולי העברת אותות טבעיים. <sup> 3 בפרט, אנו מאמינים כי על ידי עיצוב סוכנים סינטטיים המחקים את תפקודם של חלבוני איתות, אפשר יהיה לווסת את הפונקציה של התא בעקיפין. לדוגמא, רשתות המלאכותיות אלה עשויות לאפשר סמנים ביולוגיים חלבון להפעיל אנזימים לבקע prodrugs. לחלופין, mimetics חלבון איתות אלה יוכל להפעיל מסלולי איתות תאי טבעי, וכתוצאה מכך השפעות טיפוליות.
כדי להוכיח את ההיתכנות של גישה זו, יצרנו לאחרונה 'מתמר כימי "סינתטי 4 המאפשר גורם גדילה הנגזרות טסיות (PDGF) כדי לעורר את המחשוף של prodrug נגד סרטן על ידי הפעלת גלוטתיון-s-transferase (GST), אשר הוא לא שותף המחייב טבעי. המבנה של "מתמר" זה מורכב aptamer DNA אנטי PDGF כי הוא שונה עם מעכב הדו-ערכי עבור GST. לפיכך, סוכן סינתטי זה שייך למשפחה של מולקולות עם אתרי הקישור כדיחלבונים שונים, 5-7 כגון מעוררים כימיים של dimerization (מספרי CID) 8-10 וגם לקבוצת חלבון-קלסרים מבוססת על conjugates מולקולה סינטטי oligonucleotide. 11-21
העקרונות הכלליים שבבסיס העיצוב של מערכות כאלה מתוארים במסמך זה ו פרוטוקולים מפורטים סינתזה ובדיקת התפקוד של זה 'מתמר' עם מבחני האנזימטית קונבנציונליים מסופקים. עבודה זו נועדה להקל על פיתוח 'מתמרים' נוסף של מחלקה זו, אשר עשוי לשמש לתווך תקשורת בין חלבונים תאיים וכתוצאה מכך, לגרום מסלולי איתות תאים מלאכותיים.
איור 1 סכמטי המתאר את עקרונות ההפעלה של הסינתטי "מתמר כימי 'שיכול לתווך תקשורת בין חלבונים טבעיים. באיור זה, א 'מתמר כימית', משלבת קלסרים סינטטי protEins I ו- II (קלסרים I ו- II), מאפשרים חלבון השני כדי לעורר את הפעילות הקטליטית של חלבון לי, שהוא לא שותף המחייב טבעי. בהיעדר חלבון II, מתמר נקשר לאתר הקטליטית של האנזים (אני חלבון) ומעכב את פעילותו (איור 1, המדינה השנייה). הכריכה של "מתמר" לחלבון השני, לעומת זאת, מקדם אינטראקציות בין לי קלסר ועל פני השטח של חלבון II (איור 1, מדינת iii), אשר מפחית זיקה כלפי I. חלבון כתוצאה מכך, הוא הריכוז היעיל של ' מתמר חינם 'בפתרון הצטמצם, מה שמוביל דיסוציאציה של החלבון מתמר שאני מורכב וכדי מחדש של חלבון אני (איור 1, מדינת iv). יחדיו, את הפעולות הבאות להדגיש שלושה עקרונות יסוד שביסוד העיצוב של 'מתמר' יעיל: (1) "מתמר" צריך קלסר ספציפי עבור כל אחת מן מטרות החלבון, (2) betwe האינטראקציהen קלסר השני והחלבון שני צריכים להיות חזק יותר את האינטראקציה בין לי קלסר והחלבון לי, ו- (3) קלסר אני חייב להיות מסוגל לקיים אינטראקציה עם פני השטח של חלבון II. העיקרון אחרון זה לא בהכרח דורש קלסר שאני לבד תהיה זיקה גבוהה סלקטיביות לכיוון החלבון השני. במקום זאת, היא מבוססת על המחקרים עדכניים שלנו שהראו כי הבאת מולקולה סינטתית בסמיכות חלבון עשויה לקדם אינטראקציות בין מולקולה זו ועל פני השטח של החלבון. 19,22,23
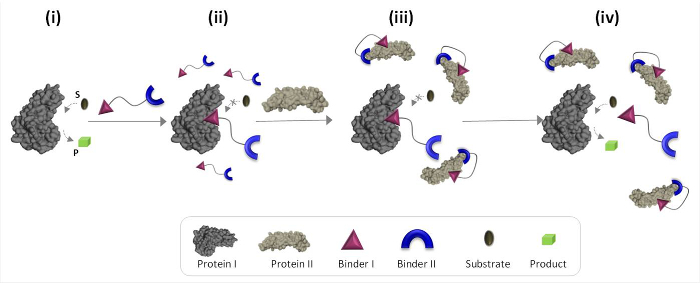
איור 1:. הפעלת עקרונות 'מתמר כימי' כאשר 'מתמר הכימית' מתווסף חלבון פעיל אני (מדינת i), הוא נקשר לאתר הפעיל שלה באמצעות קלסר לי ומעכב את פעילותו (המדינה השנייה). בנוכחות החלבון השני, לעומת זאת, לא כרוך לא כימיתransducer 'אינטראקציה עם חלבון II באמצעות קלסר השנייה, המקדמת אינטראקציות בין לי קלסר ועל פני השטח של חלבון II. קלסר המושרה זה I-חלבון II אינטראקציה מפחית את ריכוז יעיל של קלסר לי, מה שמוביל דיסוציאציה של שנות ה-חלבון transducer' שאני מורכבים לחלבון שאני מחדש (המדינה iv). נא ללחוץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו .
