Signaltransduksjonsveier spille en betydelig rolle i praktisk talt alle cellulære prosessen og tillate cellen å reagere raskt på miljøsignaler. 1. Disse banene ofte utløses ved binding av et signalmolekyl til en ekstracellulær reseptor, noe som resulterer i aktivering av intracellulære enzymer. Amplifisering og forplantning av dette signal i cellen er formidlet ved funksjon av signaleringsproteiner som danner et nettverk av protein-protein interaksjoner hvori enzymene er reversibelt aktivert med høy spesifisitet. På grunn dysregulering av disse nettverkene ofte fører til kreftutvikling, har det vært stor interesse for å etablere 'signaltransduksjon terapi av kreft', 2 hvorved medikamenter er utviklet for å forstyrre maligne signalveier. Vi har nylig foreslått en alternativ tilnærming til signaltransduksjon terapi som er avhengig av evnen til medikamenter for å generere unaturlige signaltransduksjonsveier. <sup> 3 I særdeleshet, tror vi at ved å utforme syntetiske stoffer som etterligner funksjon av signaliseringsproteiner, ville det være mulig å modulere cellens funksjon indirekte. For eksempel kan disse kunstige nettverk gjør det mulig protein biomarkører for å aktivere enzymer som spalter forløpere. Alternativt kan disse signal protein-mimetika være i stand til å aktivere unaturlige cellesignalbaner, noe som resulterer i terapeutiske effekter.
For å demonstrere gjennomførbarheten av denne tilnærmingen, har vi nylig opprettet et syntetisk "kjemiske transduser '4 som gjør det mulig blodplateavledet vekstfaktor (PDGF) for å utløse den spaltning av et anticancer prodrug ved aktivering av glutation-s-transferase (GST), som er ikke sin naturlige bindende partner. Strukturen av denne "transduser" består av et anti-PDGF-DNA aptamer som er modifisert med en bivalent inhibitor for GST. Således tilhører denne syntesemiddel til en familie av molekyler med bindingsseter for åforskjellige proteiner, 5-7 eksempel kjemiske indusere av dimerization (KBS) 8-10 og også til gruppen av protein-bindemidler basert på oligonukleotid-syntetisk molekyl konjugater. 11-21
De generelle prinsipper som ligger til grunn for utformingen av slike systemer er beskrevet her, og detaljerte protokoller for syntetisering og teste funksjonen av denne "transduser" med konvensjonelle enzymatiske analysene er gitt. Dette arbeidet er ment å forenkle utviklingen av flere transdusere '' av denne klasse som kan brukes til å formidle intracellulært protein-protein-kommunikasjon og følgelig, for å indusere kunstig celle signalveier.
Figur 1 beskriver skjematisk drifts prinsippene for syntetisk 'kjemiske transdusere "som kan megle unaturlig protein-protein kommunikasjon. I denne illustrasjonen et "kjemisk transduser ', som integrerer syntetiske bindemidler for proteins I og II (bindemidler I og II), gjør det mulig for protein II for å utløse den katalytiske aktivitet av protein I, som ikke er dens naturlig bindingspartner. I fravær av protein II, binder transduceren det katalytiske sete i enzymet (protein I) og inhiberer dets aktivitet (figur 1, tilstand ii). Bindingen av den "transduser" til protein II, men fremmer interaksjon mellom bindemiddel I og overflaten av proteinet II (figur 1, tilstand iii), noe som reduserer dens affinitet overfor protein I. Som et resultat av den effektive konsentrasjonen av ' fri 'transduser i oppløsningen er redusert, noe som fører til dissosiasjon av transduseren-protein i-komplekset og til reaktivering av protein i (Figur 1, tilstand iv). Samlet utgjør disse trinnene frem tre grunnleggende prinsipper for utforming av effektive 'transdusere': (1) en "svinger" skal ha en bestemt bindemiddel for hver av protein mål, (2) samspillet betweno bindemiddel II og protein II skal være sterkere enn vekselvirkningen mellom bindemiddel I og protein I, og (3) bindemiddel I må være i stand til å samhandle med overflaten av proteinet II. Dette siste prinsippet ikke nødvendigvis krever at bindemidlet jeg alene ville ha en høy affinitet og selektivitet overfor protein II. I stedet er det basert på vår nylige studier som viste at bringe et syntetisk molekyl i tilknytning til et protein som er egnet til å fremme interaksjon mellom dette molekylet og overflaten av proteinet. 19,22,23
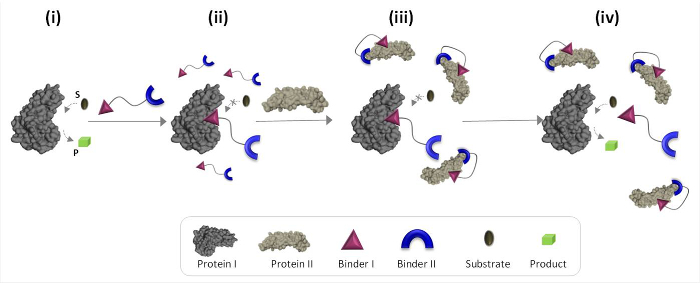
Fig. 1: Drifts prinsipper 'kjemiske transdusere' Når den "kjemiske transduseren 'tilsettes til et aktivt protein I (tilstand i), bindes det til dets aktive sete gjennom bindemiddel I og inhiberer dets aktivitet (tilstand II). I nærvær av protein II, men det ubundne 'kjemisk transducer 'samvirker med protein II gjennom bindemiddel II, som fremmer interaksjon mellom bindemiddel I og overflaten av proteinet II. Dette indusert bindemiddel I-protein II interaksjon reduserer den effektive konsentrasjonen av bindemiddel jeg, noe som fører til dissosiasjon av "transducer'-protein jeg kompleks og protein jeg reaktive (stat iv). Klikk her for å se en større versjon av dette tallet .
