This work describes the comparison of two routes of administration of BCG: subcutaneous and intranasal. Subcutaneous route is comparable to the intradermal, which is the current clinical route for BCG worldwide. Intranasal route of vaccination aims to mimic the natural route of infection of M. tuberculosis, with the objective to induce immune response directly in the lungs, the primary target organ of this pathogen.
Figure 1 describes the workflow followed. Eight to ten week-old female DBA/2 mice are vaccinated with 106 CFU of BCG Danish by the subcutaneous or intranasal route of administration. Eight weeks later, a group of mice is sacrificed to analyze lung immune response induced by vaccination. BAL samples are first obtained and then we harvest lungs. In order to study vaccine-conferred protective efficacy, we inoculate a different group of mice with a low-dose intranasal challenge of M. tuberculosis H37Rv strain. One month later, we sacrifice animals and harvest lungs. In this case, for each animal we use the left lung to determine bacterial load and the right lung to assess vaccine-induced immune response post-challenge. The objective is to generate bacterial load and immune response data in lungs for each animal in order to study potential correlates of protection.
As shown in Figure 2, lungs were dissociated to obtain either an organ homogenate or a cellular suspension. Homogenized lungs were plated on solid agar medium to determine bacterial load four weeks post-challenge. Cellular suspension to study vaccine-induced immune response was obtained following lung enzymatic digestion with collagenase D and DNaseI.
Our results clearly indicate that, when compared to the subcutaneous route, the intranasal route of vaccination confers a much greater protective efficacy in lungs four weeks after challenge (Figure 3A). In addition, we confirmed that bacterial load in lungs from the intranasal BCG-vaccine group correspond to H37Rv and not to BCG vaccine. To this end, we analyzed a representative number of colonies from this group by specific PCR for the RD9 genome region, which amplifies different length fragments in BCG and M. tuberculosis (Figure 3B). Figure clearly showed that all the colonies analyzed provided a fragment of 0.4 kbp, which corresponded to H37Rv.
Our data revealed correlation between protective efficacy conferred by intranasal BCG vaccination and vaccine-induced immune response in lungs prior to challenge. Intranasal BCG clearly triggered higher IL17 and IFNγ production in lungs, measured by ELISA (Figure 4A). These data were confirmed by intracellular staining (ICS) and flow cytometry (data not shown). In addition, we also found a higher concentration of both total and PPD-specific IgA concentration in BAL samples (Figure 4B), indicating that pulmonary BCG vaccination induces production of IgA and translocation to respiratory airways.
Finally, we studied BCG-induced immune response in lungs after challenge (Figure 5). Our data revealed differences between IL17 and IFNγ; IL17A-producing CD4+ cells were only detected in the intranasal BCG group, whereas IFNγ-producing cells were found in all groups infected with H37Rv regardless of vaccination. Representation of data of each animal corresponding to IL17-producing cells and bacterial load showed a significant correlation between presence of IL17 and lung bacterial load reduction, which was not observed in the case of IFNγ.
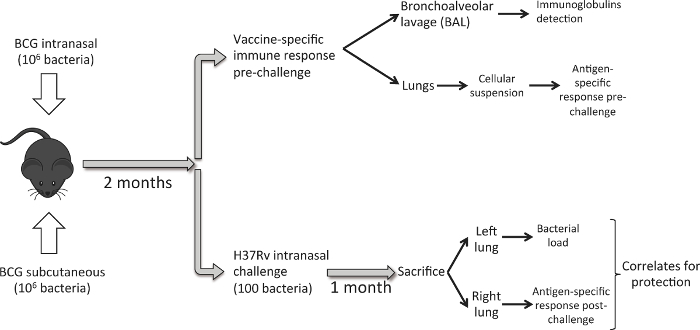
Figure 1. Workflow to Compare Intranasal and Subcutaneous Route of BCG Vaccination. Eight weeks post BCG immunization, a set of mice (6 per experimental group) is used to harvest lungs and perform a BAL, and analyze pulmonary immune response induced by vaccination. Another set of mice (6/group), is inoculated with a low-dose intranasal challenge of M. tuberculosis H37Rv strain (100 CFU). One month later, animals are sacrificed and bacterial load is analyzed in the left lung and PPD-specific immune response in the right lung. Please click here to view a larger version of this figure.
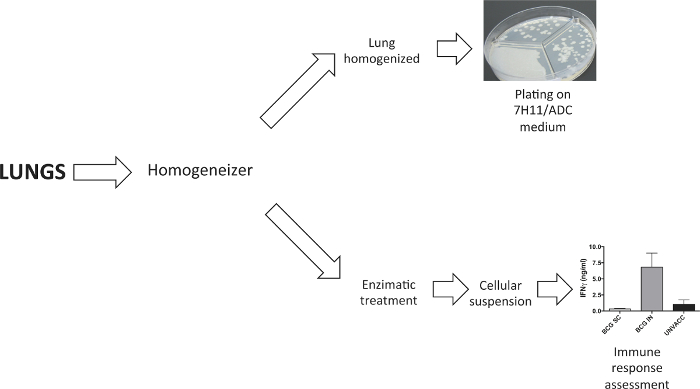
Figure 2. Processing of Lung Samples. A tissue dissociator was used to process lungs. In the experiments to determine bacterial load, lungs were homogenised prior to plate them in solid agar medium. In experiments that require a lung cellular suspension, this was generated following enzymatic digestion with collagenase D and DNaseI. Please click here to view a larger version of this figure.
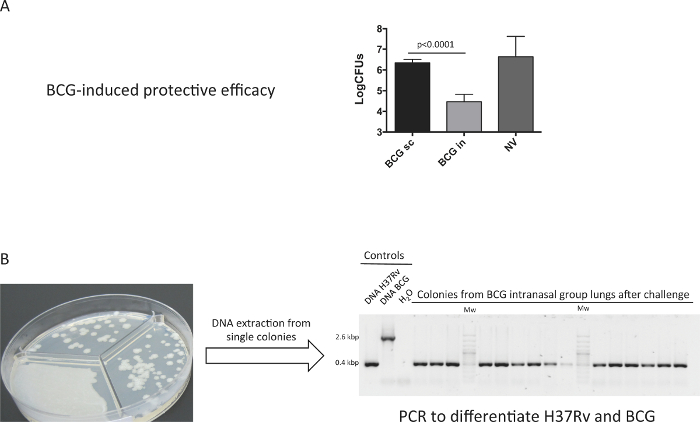
Figure 3. Protective Efficacy Conferred by BCG Immunization. Groups of 6 DBA/2 mice were vaccinated by the subcutaneous (BCG sc), intranasal (BCG in) route, or non-vaccinated (Unvacc) with BCG Danish vaccine 106 CFU. At two months post-vaccination, mice were inoculated intranasally with a low dose (100 CFU) H37Rv challenge, and one-month later bacterial burden in lungs was determined. A representative experiment of two independent is shown. (A) Data in the graphs are represented as mean+SD. One-way ANOVA test with Bonferroni post analysis was performed to calculate statistical significance. (B) A representative number of single colonies from the BCG intranasal group were analyzed by PCR specific for RD9 region (different in BCG and H37Rv genomes) to discern BCG and H37Rv colonies. (Previously published 9). Please click here to view a larger version of this figure.
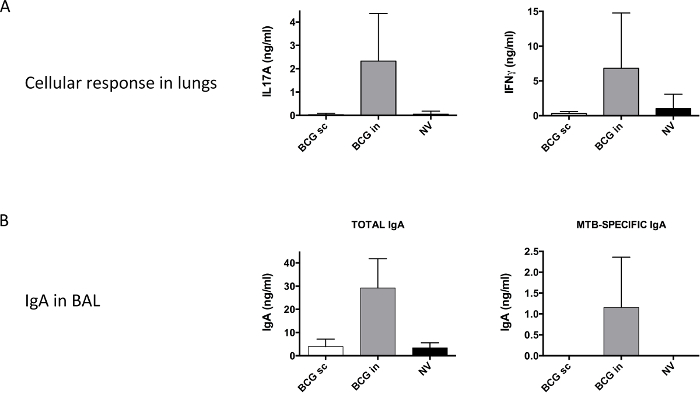
Figure 4. Vaccine-specific Pulmonary Immune Response Analyzed Prior to Challenge with H37Rv. Groups of 6 DBA/2 mice were vaccinated by the subcutaneous (BCG sc) or intranasal (BCG in) route, or non-vaccinated (Unvacc) with BCG Danish vaccine 106 CFU. (A) At two months post-vaccination, a cellular suspension from harvested lungs was obtained. Cells were stimulated with PPD as described in methods section and IL17A (left panel) and IFNγ (right panel) production were analyzed by ELISA. (B) Total IgA, and M. tuberculosis (MTB)-specific IgA were analyzed from BAL samples by ELISA. Pooled data from two independent experiments are shown. Data in the graphs are represented as mean + SD. (Previously published 9). Please click here to view a larger version of this figure.
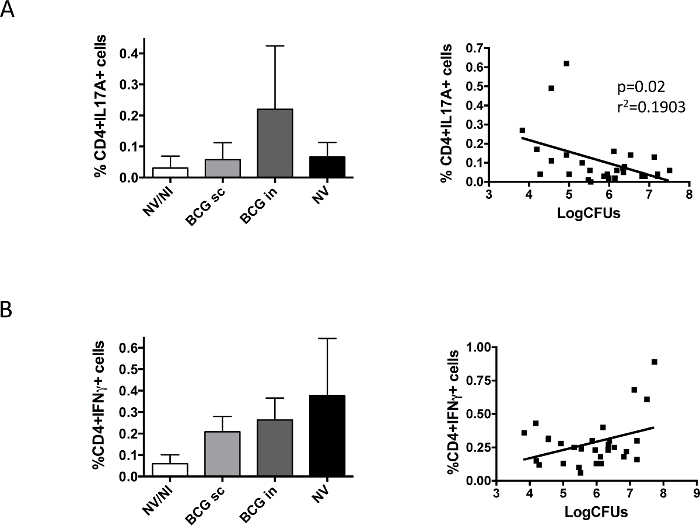
Figure 5. Vaccine-specific Pulmonary Immune Response Analyzed prior to Challenge with H37Rv. Groups of 6 DBA/2 mice were vaccinated by the subcutaneous (BCG sc) or intranasal (BCG in) route, or non-vaccinated (NV) with BCG Danish vaccine 106 CFU. A control group of non-vaccinated, non-infected mice was also included (NV/NI). At two months post-vaccination, mice were challenged intranasally with a low H37Rv dose (100 CFU), and one-month later animals were euthanized. Left and right lungs from the same animal were used to determine bacterial load and IL17A- (A) or IFNγ- (B) producing CD4+ cells, respectively. Data in left panels correspond to percentage of cytokine-producing cells measured by flow cytometry, and are represented as mean + SD. Right panels represent data from bacterial load and cytokine- producing CD4+ cells obtained for each mouse. Linear regression was calculated and the p-value obtained in each case is shown in the case of IL17A. Pooled data from two independent experiments are shown in the figure. (Previously published 9). Please click here to view a larger version of this figure.
