The above protocol allows visualization of the punctate structures associated with mitochondria (Figure 1B-C), which indicate mitochondrial DNA replication during Drosophila oogenesis. The EdU puncta localized with mitochondria marked by staining for ATP synthase alpha subunit (Figure 2). The observed signals were absent in ovaries treated with ethidium bromide 11, an inhibitor for mtDNA replication 14, validating that these puncta indeed label replicating mtDNA.Aphidicolin was used to inhibit nuclear DNA staining without affecting mtDNA replication (Figure 2). Without aphidicolin treatment, intense EdU signals label the nuclei, and mtDNA puncta were barely detected (Figure 3). However, in the presence of aphidicolin, nuclear incorporation was dramatically reduced and many puncta associated with mitochondria were observed.
There is a high level of mtDNA replication in post-germarium egg chambers (Figure 1B). However, notably, mtDNA replication displayed a spatial pattern in the germarium. As indicated by the number of EdU puncta, there is a moderate level of mtDNA replication in region 1 of the germarium, but almost no EdU incorporation in region 2A (Figure 1C). As the cyst moves down to region 2B in the germarium, mtDNA replication resumed and the number of EdU puncta in the posterior cyst of region 2B was much higher than that in region 2A (Figure 1C). Specifically, intensive EdU incorporation was concentrated around the ring canals and the fusome structures, as stained by the hu li tai shao (Hts) protein. mtDNA kept replicating at a high level in region 3 of the germarium (Figure 1C)
To demonstrate the association of specific genes or treatment with mtDNA proliferation, Drosophila ovaries could be subjected to gene manipulation or drug treatment. We treated ovaries with different concentrations of FCCP, a classic mitochondrial protonophore, which dissipates mitochondrial membrane potential. As a control, DMSO had no effect on mtDNA replication (Figure 4A). High doses of FCCP (10 µM) almost depleted mtDNA replication entirely throughout the germarium (Figure 4D). Nonetheless, lower concentration of FCCP (2 or 5 µM) had a minor impact on mtDNA replication in region 1 but inhibited replication in regions 2B and 3 (Figure 4B-C), suggesting regions 2B and 3 are more sensitive to mitochondrial disruption, or they maintain relative slower replication kinetics. The above results indicated that mtDNA replication is associated with mitochondrial activity. Particularly, different regions of the germarium responded differently to mitochondrial impairment.
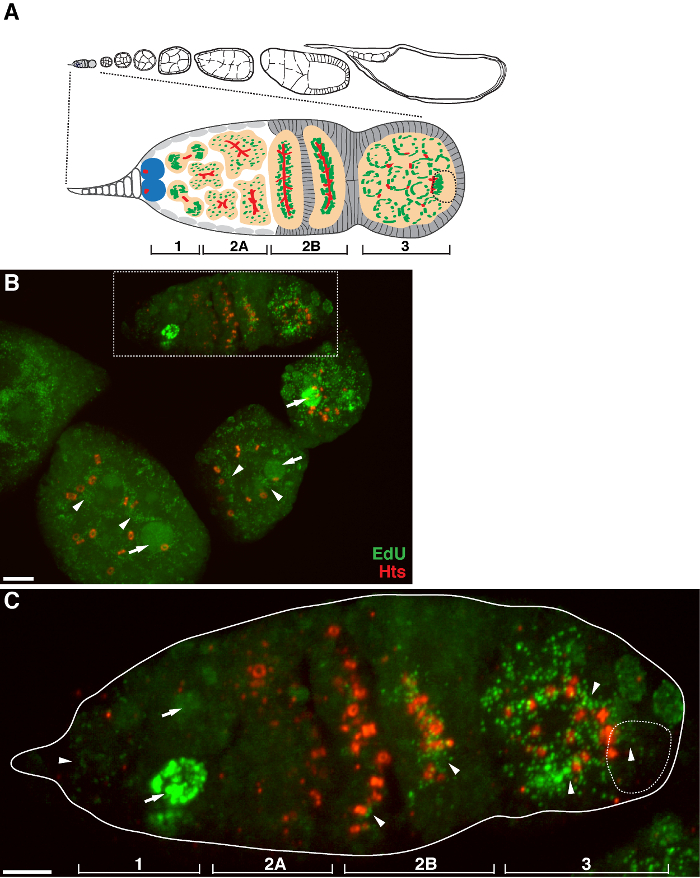
Figure 1. mtDNA replication during Drosophila oogenesis. (A) Diagram of a Drosophila ovariole and a magnified view of the germarium. The ovariole illustrates from left to right, anterior to posterior, successive developmental stages of egg chambers. In the germarium, the fusome (red), germline stem cells (blue), mitochondria (green), future oocyte (broken line, recognized by positioning, fusome structure and mitochondrial clusters), developing cysts (peach) and four developmental regions are shown. (B) Representative z-stack projection of a wild-type ovariole labeled by EdU and antibody against Hts-RC, a marker for ring canals and fusome. In the presence of aphidicolin, the EdU was incorporated into mtDNA (arrowheads) and nuclei (arrows). Scale bar, 10 µm. (C) Magnified view of germaria outlined in the boxed region in B. The four developmental regions are indicated. Scale bar, 5 µm 11 . Please click here to view a larger version of this figure.
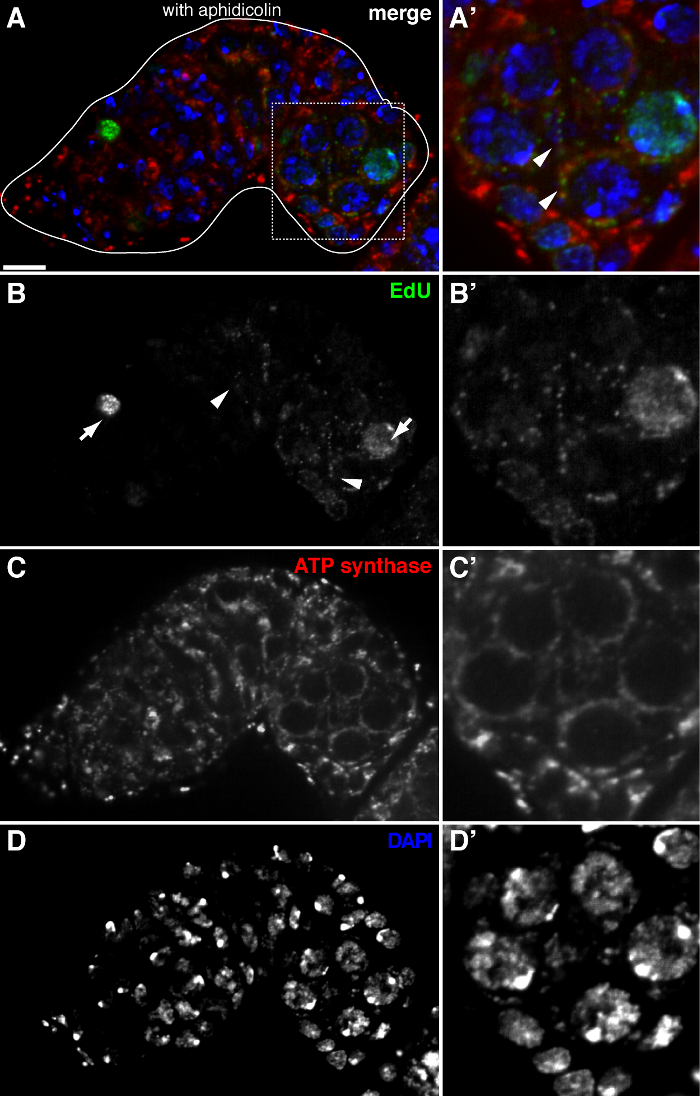
Figure 2. mtDNA replication in the Drosophila germarium visualized by EdU incorporation with aphidicolin treatment. (A)-(D) Representative confocal section of a wild type germarium showing EdU incorporation (green, b), mitochondria, marked by ATP Synthase alpha subunit staining (red, C), and nuclei, labeled with DAPI staining (blue, D) with pre-incubation with DNA polymerase-α inhibitor aphidicolin. (A'-D') A magnified image of the boxed area in (A) showing EdU incorporation (B'), mitochondria (C') and nuclei (D'). In the presence of aphidicolin, nuclear incorporation (arrows) is reduced and many puncta were localized within mitochondria (arrowheads). Scale bars, 10 µm. The figure has been modified from 11. Please click here to view a larger version of this figure.
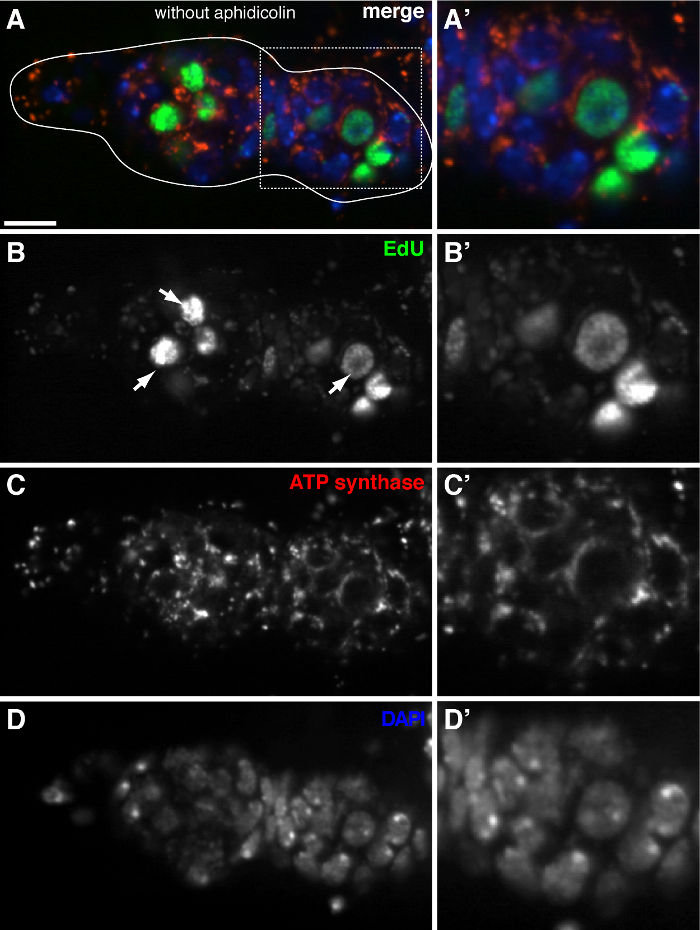
Figure 3. mtDNA replication is barely detected in the Drosophila germarium without aphidicolin treatment. (A)-(D) Representative confocal section of a wild type germarium showing EdU incorporation (green), mitochondria, marked by ATP Synthase alpha subunit staining (red), and nuclei, labeled with DAPI staining (blue) in the absence of the DNA polymerase-α inhibitor aphidicolin. (A'-D') A magnified image of the boxed area in (A) showing EdU incorporation (B'), mitochondria (C') and nuclei (D'). Without aphidicolin, intense EdU signals label nuclei (arrows), and mtDNA puncta were barely detected. Scale bars, 10 µm. The figure has been modified from 11. Please click here to view a larger version of this figure.
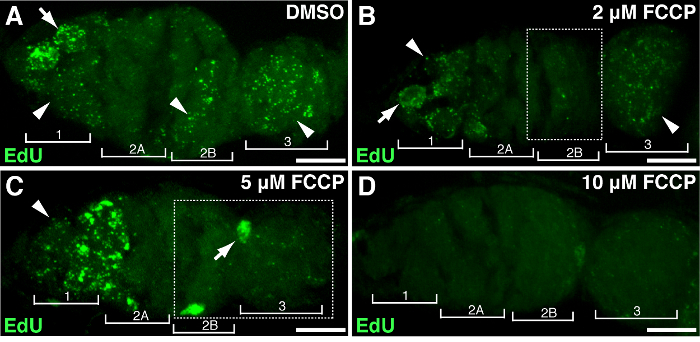
Figure 4. Mitochondrial uncoupler impairs the mtDNA replication. Representative z-stack projects showing wild-type germarium treated with DMSO (A) or the mitochondrial uncoupler FCCP at concentrations of 2 µM (B), 5 µM (C), 10 µM (D) during EdU incorporation. Note the impaired EdU labeling in region 2B treated with 2 µM FCCP (B), and in both region 2B and 3 treated with 5 µM FCCP (outlined in boxes). Four developmental regions are shown. Arrows, nuclear DNA; arrowheads, mtDNA. Scale bar, 10 µm. The figure has been modified from 11. Please click here to view a larger version of this figure.
