Polymorfonukleære neutrofiler (PMN) udgør den største befolkning af leukocytter i blodet, der tjener som den første linje i forsvaret mod invaderende patogener ved at producere en bred vifte af cytotoksiske molekyler. Den traditionelle opfattelse har længe været, at af blod cirkulerer, kortlivet, professionelle fagocytter, som er de første til at ankomme til akutte inflammatoriske steder til at bekæmpe infektioner og støtte i clearance af patogener og skadelige partikler. 1 I deres ikke-aktiveret tilstand, neutrofiler er konstitutivt engageret i apoptose. Når du overfører fra blodet til inflammatoriske steder, neutrofiler gennemgår aktivering til at løse inflammation. De fagocytere og dræbe invaderende mikroorganismer, ved at producere en bred vifte af cytotoksiske molekyler som reaktive ilt arter (ROS), lytiske enzymer såsom neutrofilelastase (NE) og cathepsiner med potent mikrobicid aktivitet. For at fælde patogener, neutrofiler også frigive ekstracellulære fælder (NET), Der består af nukleare kromatin tråde indeholder antibakterielle peptider og forskellige lytiske enzymer. Dog kan ukontrolleret udslip af disse cytotoksiske molekyler fra neutrofiler også forevige inflammatoriske reaktioner og fremkalde skader på omgivende væv. 2 Derfor er en effektiv clearance af apoptotiske neutrofiler af makrofager (MO) og dendritiske celler (DC) er afgørende for at løse inflammation. 3, 4, 5, 6
I de senere år er der imidlertid er det blevet mere og mere klart, at neutrofiler er alsidige celler, hvis funktioner går langt ud over fagocytose og patogen drab. 6, 7 Ved undergår priming eller aktivering, er neutrofil plasticitet langsomt ved at vinde opmærksomhed. For eksempel bakterier og mycobakterier udfordret neutrofiler blev vist tiludskiller interleukin (IL) -10 og styre den inflammatoriske reaktion, hvilket antyder tilstedeværelsen af immuno-regulatoriske responser. 8 Post-mitotiske neutrofiler blev vist til trans-differentiere til MO-lignende celler, eller DC-lignende celler ved fordøjelse og præsentere antigen fragmenter, når de behandles med cytokiner og vækstfaktorer, 9, 10 således, betjener en kritisk rolle i integrationen af medfødte og adaptive reaktioner. 3, 6 Aktivering af vækstfaktorer fremmet engulfment af apoptotiske neutrofiler eller cellerester derved lette clearance af vragdele på inflammatoriske steder og løsningen af inflammation, 3, 9 især når MO / DC clearance system er utilstrækkelig eller overvældet, 11, 12 tyder potentielle "selvregulering" for at hjælpe reløse det inflammatoriske respons. Dette, idet apoptose er en form for reguleret selv-dødsfald, som kan inhibere ekstracellulær frigivelse af cytotoksiske forbindelser og dermed forhindre skade på omgivende væv. 6
Langvarig overlevelse er en anden funktion af neutrofil aktivering og blev påvist ved behandling med forskellige vært afledte faktorer såsom granulocytkolonistimulerende faktor (G-CSF), granulocyt-makrofag koloni-stimulerende faktor (GM-CSF), inflammatoriske cytokiner, såsom interferon ( IFN) -γ, tumornekrosefaktor (TNF) -a og / eller patogen afledte produkter, hvori der tages neutrofiler til at modulere deres overlevelse respons. 6 Faktisk neutrofil overlevelse er en forudsætning for dets plasticitet og var forbundet med dens evne til at udføre fagocytose. 6, 13 Følgelig blev det også vist at associere med fænotypiske og funktionelle ændringer, som DEPENded på opreguleret genekspression ved at inducere syntese af nye proteiner involveret i neutrofil levetid forlængelse, og formindsket apoptose. 10
I modsætning til neutrofiler, som er kortvarig og konstitutivt gennemgår apoptose i kultur, eller de cytokiner / vækstfaktorer-aktiverede neutrofiler, der er beskrevet ovenfor, som har forlænget levetid, har vi for nylig identificeret en ny, lille delpopulation af neutrofiler, der udvikler spontant i langvarig standard kultur betingelser fra frisk isolerede humane blodneutrofiler uden eksternt tilsætning cytokiner eller vækstfaktorer. 14 Disse neutrofil-afledte celler, som ikke var beskrevet før i litteraturen blev betegnet giant fagocytter (Gφ). Den Gφ har udvidet levetid i kultur, de er fuldt udviklet inden for 5-7 dage, og er kendetegnet ved unikke morfologiske træk, fænotypiske udtryk og funktioner. De er voldsomt forstørret på grund af autophagocytosis af døde neutrofile rester, vakuoliserede, og indeholder phagolysosomes. Den Gφ udtrykke specifikke neutrofile granula markør – cluster of differentiation (CD) 66b, de azurofile granuler markører – CD63 og myeloperoxidase (MPO) og yderligere neutrofile markører, såsom CD11, NE, CD15, de NADPH oxidase subunits gp91- phOx og p22 phOx og autophagy markør -LC3BII. 14, 15 Funktionelt de aktivt take-up latexkugler og zymosan partikler, og generere ROS i afhængighed zymosan og phorbol-12-myristat-13-acetat (PMA) stimulation. Interessant, i modsætning til friske neutrofiler, Gφ også intensivt udtrykke scavenger receptorer CD68 og CD36, take-up oxideret low density lipoprotein (oxLDL), og generere ROS i respons på stimulering med oxLDL. Derudover Gφ er blottet for de monocytiske afstamning markører CD14, CD16 og CD163 eller de dendritiske markører CD1c og CD141. Desuden PHAgocytosis og autophagy og sandsynligvis funktionelle NADPH oxidase er forudsætninger for deres udvikling. Dette eftersom, fagocytose-inhibitor cytochalsin B, den autofagi inhibitorer 3-methyladenin (3-MA) og bafilomycin (BafA1) og NADPH oxidase inhibitor – diphenylen iodonium (DPI) – forhindrede deres udvikling. Derudover monocytter / neutrofiler co-kulturer samt udsættelse for intermitterende hypoxi hæmmet deres udvikling, mens neutrofil tilpasning til vedvarende hypoxi var tydelig. 14,15 Deres foreslåede udvikling i kultur er illustreret i figur 1 .Den protokol i nærværende dokument beskriver trin for trin forberedelse af Gφ fra frisk isoleret cirkulerende humant blod neutrofiler, deres udvikling, identifikation og nogle grundlæggende karakteristika. Denne protokol kan bruges til at undersøge og afsløre det brede spektrum og roller disse nyligt beskrevet og spændende neutrofil-afledt Gφ for at characterize deres betydning og deres potentielle funktioner.
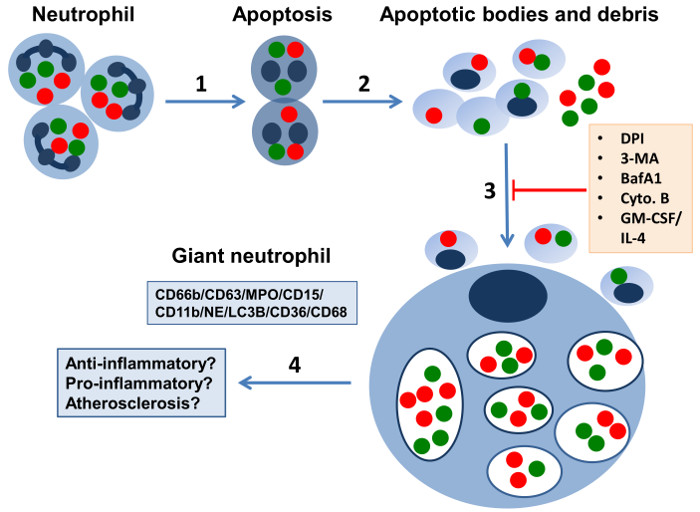
Figur 1: Skematisk repræsentation kæmpeceller Udvikling i 7 Day Neutrocyt kulturer. Det foreslås, at der på inflammatoriske steder (1) neutrofiler undergår apoptotisk celledød, og (2) release-membran-omkranset fragmenter indeholdende nukleart affald, granulat (grønne og røde prikker) og andre subcellulære bestanddele, som udløser autofagi mekanismer. (3) Giant fagocytter (Gφ) udvikle sig i langsigtede neutrofile kulturer blottet for cytokiner eller vækstfaktorer ved internalisere apoptotiske organer og neutrofil vragrester, samtidig med at funktionelle NADPH oxidase.They er kendetegnet ved forskellige neutrofil CD66b + / CD63 + / MPO + / CD15 + / CD11 + / NE markører, store fagosomer omslutter granulater og cellerester, og scavenger receptorer CD36 og CD68. Gb1; er for det meste mononukleære celler, i stand til at internalisere også forskellige partikler og oxideret LDL og generere ROS. Membranerne af celleblærerne påfyldning Gφ indeholder LC3B (markeret med mørkeblå), en markør for autophagosomal membran, hvilket tyder på en streng sammenhæng mellem autofagi og kæmpe fagocyt formation. Gφ ikke udvikler i medium indeholdende GM-CSF / IL-4. Også hæmmere såsom NADPH oxidase inhibitor – diphenylen iodonium (DPI), de autofagi hæmmere 3-methyladenin (3-MA) og bafilomycin (BafA1) og fagocytose inhibitor cytochalasin B (. Cyto B) afskaffe deres dannelse. (4) Potentielle Gφ funktioner i vivo kan omfatte anti eller pro-inflammatoriske egenskaber og deltagelse i aterosklerotiske processer (dette tal er baseret på vores resultater 14, 15 og blev ændret fra den medfølgende Leder af Berton 20). Klik her for at se en større version af dette tal.
