Полиморфноядерные нейтрофилы (ПМН) составляют самую большую популяцию лейкоцитов в крови, выступающей в качестве первой линии обороны против вторжения патогенных микроорганизмов, производя широкий спектр цитотоксических молекул. Традиционная точка зрения уже давно, что профессиональных фагоцитов крови, циркулирующей непродолжительным, которые являются первыми, чтобы прибыть в острых воспалительных очагов бороться с инфекциями и помощь в зазоре патогенов и вредных частиц. 1 В их неактивированной состоянии, нейтрофилы конститутивно приверженность к апоптозу. При миграции из крови в местах воспаления, нейтрофилы подвергаются активации для разрешения воспаления. Они фагоцитируют и убить вторжения микроорганизмов, производя множество цитотоксических молекул в качестве активных форм кислорода (ROS), литические ферменты, такие как эластазы нейтрофилов (NE) и катепсинов с мощной бактерицидной активностью. Для того, чтобы улавливать патогенами, нейтрофилы также выделяют внеклеточные ловушки (сетки), Которые состоят из ядерного хроматина нитей, содержащих антибактериальные пептиды и различные литические ферменты. Тем не менее, неконтролируемый выброс этих цитотоксических молекул из нейтрофилов может также увековечить воспалительные реакции и вызывают повреждение окружающих тканей. 2 Таким образом, эффективный клиренс апоптотических нейтрофилов макрофагами (М ^) и дендритные клетки (DC) имеет решающее значение для устранения воспаления. 3, 4, 5, 6
В последние годы, однако, становится все более очевидным, что нейтрофилы являются весьма разносторонние клетки, функции которых выходят далеко за пределы фагоцитоза и патогена убийства. 6, 7 Соблюдая гигиену грунтование или активацию нейтрофилов пластичностью постепенно завоевывает все внимание. Например, бактерии и микобактерии оспаривались нейтрофилы показаносекретируют интерлейкин (IL) -10 и контролировать воспалительную реакцию, что указывает на наличие иммуно-регуляторных реакций. 8 постмитотическими нейтрофилы показано транс-дифференцироваться в М ^-подобных клеток, или постоянного тока , таких как клетки при переваривании и представляя антиген фрагментов при обработке цитокинов и факторов роста, 9, 10 , таким образом, служит решающую роль в интеграции врожденной и адаптивной ответы. 3, 6 Активация факторов роста способствовали при полном охвате апоптотических нейтрофилов или остатков клеток, тем самым, облегчая очистку мусора в местах воспаления и разрешение воспаления, 3, 9 в частности , когда система зазора М ^ / DC является недостаточным или перегружены, 11, 12 предлагая потенциал "саморегулирование", чтобы помочь восстановитьрешить воспалительную реакцию. Это, так как апоптоз является формой регулируемой собственной смерти, которое может ингибировать внеклеточный высвобождение цитотоксических соединений, и тем самым предотвратить повреждение окружающих тканей. 6
Увеличивал выживаемость является еще одним признаком активации нейтрофилов и была продемонстрирована путем обработки различными принимающими получены факторы, такие как гранулоцитарный колониестимулирующий фактор (G-CSF), гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), воспалительных цитокинов, таких как интерферон ( ИФН) -γ, фактора некроза опухоли (ФНО) и / или патогена, полученных продуктов, таким образом, позволяя нейтрофилы модулировать их реакцию выживания. 6 На самом деле, выживание нейтрофилов является необходимым условием для его пластичности и было связано с его способностью выполнять фагоцитоз. 6, 13 Соответственно, было также показано , что ассоциируют с фенотипическими и функциональными изменениями , которые висимостьDed на повышающей регуляции экспрессии генов, индуцируя синтез новых белков, участвующих в нейтрофильных расширении продолжительность жизни, и уменьшалась апоптоза. 10
В отличие от нейтрофилов, которые недолго и конститутивно апоптозу в культуре, или цитокины / факторы роста активированные нейтрофилы, описанные выше, которые продлевал срок службы, в последнее время мы идентифицировали новую, небольшой субпопуляции нейтрофилов, которая развивается спонтанно в продолжительном стандартной культуре условия из свежеизолированных нейтрофилов крови человека без добавления извне цитокины или факторы роста. 14 Эти нейтрофильных полученные клетки, которые не были описаны ранее в литературе были названы гигантские фагоциты (Сф). Сф продлили продолжительность жизни в культуре, они полностью разработаны в течение 5-7 дней, и характеризуются уникальными морфологическими признаками, экспрессии фенотипического и функций. Они значительно увеличены за счет autophagoЦитоз мертвых остатков нейтрофильных, вакуолизированных, и содержат фаголизосомах. Сф выражают специфический маркер гранул нейтрофилов – кластер дифференцировки (CD) 66b, азурофильных гранулы маркеров – CD63 и миелопероксидазы (МРО) и дополнительные нейтрофилы маркеры , такие как CD11b, NE, CD15, НАДФН – оксидазы субъединиц gp91- phox и p22- phox и аутофагия маркер -LC3BII. 14, 15 Функционально, они активно принимают меры латексные шарики и частицы зимозаном, а также генерировать ROS в ответ на зимозаном и форбол 12-миристат 13-ацетата (РМА) стимуляции. Интересно, что в отличие от свежих нейтрофилы, Сф также интенсивно выражают мусорщик рецепторов CD68 и CD36, натяжные окисляется липопротеинов низкой плотности (oxLDL), а также генерировать ROS в ответ на стимуляцию oxLDL. Кроме того, Сф лишены моноцитарных маркеров CD14 линии дифференцировки, CD16 и CD163 или дендритные маркеры CD1c и CD141. Кроме того, PHAgocytosis и аутофагии и, вероятно, функциональная NADPH оксидазы являются необходимыми условиями для их развития. Это с тех пор, фагоцитоза-ингибитор cytochalsin В, ингибиторы автофагии 3-метиладенин (3-МА) и bafilomycin (BafA1) и ингибитор оксидазы NADPH – дифенилен иодония (DPI) – предотвратить их развитие. Кроме того, моноциты / нейтрофилы сокультурах, а также воздействие прерывистой гипоксии препятствует их развитию, в то время как нейтрофилы адаптация к устойчивой гипоксии была очевидна. 14,15 Их развитие в Рекомендованное культуры показана на рисунке 1 протокол .The в настоящем документе описывается шаг за шагом подготовки от G ^ свежевыделенные циркулирующих нейтрофилов крови человека, их развитие, идентификацию и некоторые основные характеристики. Этот протокол может быть использован для дальнейшего исследования и выявить широкий спектр и роль этих недавно описал и интригующим нейтрофилы происхождения Сф для того, чтобы characterizе их значение и их потенциальные функции.
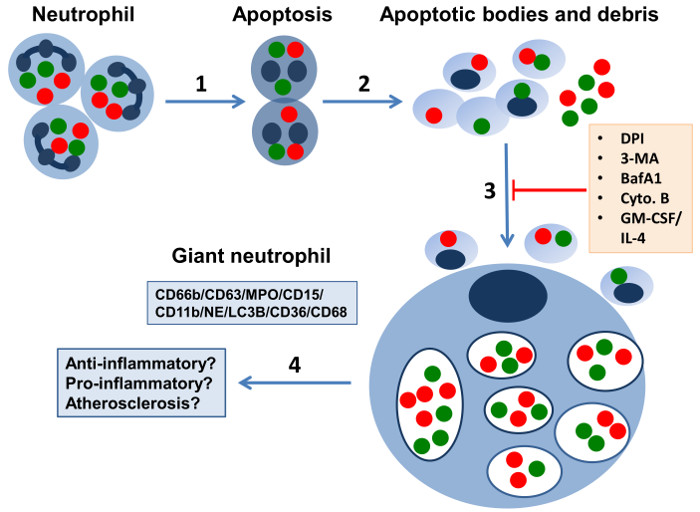
Рисунок 1: Схематическое изображение развития гигантских клеток в 7 -й день нейтрофильных культур. Предполагается , что на участках воспаления (1) нейтрофилы подвергаются апоптотической гибели клеток, и (2) релиз мембраной окружении фрагментов , содержащих ядерный мусор, гранулы (зеленые и красные точки) и другие субклеточные компоненты , которые запускают механизмы аутофагии. (3) Гигантские фагоциты (Сф) развиваются в долгосрочных нейтрофильных культур , лишенных цитокинов или факторов роста усваивая апоптотических телец и нейтрофилов мусора, сохраняя при этом функциональный NADPH oxidase.They характеризуются различными нейтрофильного CD66b + / CD63 + / MPO + / CD15 + / CD11b + / NE маркеры, большие фагосомы ограждающих гранул и остатков клеток и мусорщик рецепторов CD36 и CD68. гигабайт1; в основном мононуклеарные клетки, способные к интернализации также различные частицы и окисленного LDL и генерировать ROS. Мембраны вакуолей заполнения Сф содержат LC3B (отмечены темно-синим цветом), маркера autophagosomal мембраны, что предполагает строгую ассоциацию между аутофагии и формирования гигантского фагоцитов. Сф не развиваются в среде, содержащей GM-CSF / IL-4. Кроме того, ингибиторы, такие как ингибитор NADPH-оксидазы – дифенилен иодония (DPI), ингибиторы автофагии 3-метиладенин (3-МА) и bafilomycin (BafA1) и ингибитор фагоцитоз цитохалазином (. Цито B) отменить их образование. (4) Потенциальные функции Сф в естественных условиях может включать в себя анти- или провоспалительные свойства и участие в атеросклеротических процессах (эта цифра основана на наших выводах 14, 15 и был изменен с сопровождающим редакционную BЭртон 20). Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
