Polymorfonukleära neutrofiler (PMN) utgör den största populationen av leukocyter i blodet, som fungerar som den första försvarslinjen mot invaderande patogener genom att producera ett brett spektrum av cytotoxiska molekyler. Den traditionella synen har länge varit att cirkulerande blod, kortlivad, professionella fagocyter, som är de första att anlända till akuta inflammatoriska platser för att bekämpa infektioner och stöd i avslutandet av patogener och skadliga partiklar. 1 I sin icke-aktiverade tillstånd, neutrofiler är konstitutivt engagerade i apoptos. När du migrerar från blodet till inflammationsställen, neutrofiler genomgå aktivering för att lösa inflammation. De fagocytera och döda invaderande mikroorganismer, genom att producera en rad cytotoxiska molekyler som reaktiva syreföreningar (ROS), lytiska enzymer såsom neutrofilelastas (NE) och katepsiner med potent mikrobicid aktivitet. För att fånga patogener, neutrofiler frisläpper också extracellulära fällor (NET) Som består av kärn kromatin trådar som innehåller antibakteriella peptider och olika lytiska enzymer. Emellertid kan okontrollerat utsläpp av dessa cytotoxiska molekyler från neutrofiler också föreviga inflammatoriska svar och framkalla skador på omgivande vävnader. 2 Därför är en effektiv clearance av apoptotiska neutrofiler genom makrofager (M O) och dendritiska celler (DC) avgörande för att lösa inflammation. 3, 4, 5, 6
Under de senaste åren har det dock blivit allt tydligare att neutrofiler är mycket mångsidiga celler, vars funktioner går långt utöver fagocytos och patogen dödande. 6, 7 genom att genomgå priming eller aktivering är neutrofila plasticitet gradvis att få uppmärksamhet. Till exempel, bakterier och svamp utmanade neutrofiler visade sigutsöndrar interleukin (IL) -10 och kontrollera den inflammatoriska responsen, vilket tyder på förekomsten av immunreglerande svar. 8 Post-mitotiska neutrofiler visade sig trans differentiera till M O-liknande celler, eller DC-liknande celler genom att smälta och presentera antigen fragment vid behandling med cytokiner och tillväxtfaktorer, 9, 10 således, som betjänar en kritisk roll för att integrera medfödda och adaptiva svar. 3, 6 Aktivering av tillväxtfaktorer främjas omvälvning av apoptotiska neutrofiler eller cellrester, och därmed underlätta clearance av skräp vid inflammationsställen och upplösningen av inflammation, 3, 9 särskilt när M O / DC klareringssystem är otillräcklig eller överväldigad, 11, 12 vilket tyder på potential "självreglering" för att hjälpa relösa det inflammatoriska svaret. Detta eftersom apoptos är en form av reglerad själv död, som kan hämma extracellulär frisättning av cytotoxiska föreningar och sålunda förhindra skada på omgivande vävnader. 6
Förlängd överlevnad är en annan funktion av neutrofil aktivering och påvisades genom behandling med olika värd härledda faktorer såsom granulocyt-kolonistimulerande faktor (G-CSF), granulocyt-makrofag-kolonistimulerande faktor (GM-CSF), inflammatoriska cytokiner såsom interferon ( IFN) -γ, tumörnekrosfaktor (TNF) -a och / eller patogena framställda produkter, alltså, så neutrofiler att modulera deras överlevnad svar. 6 I själva verket är neutrofila överlevnad en förutsättning för dess plasticitet och i samband med dess förmåga att utföra fagocytos. 6, 13 Följaktligen var det också visat sig associera med fenotypiska och funktionella förändringar som depended på uppregleras genuttryck genom att inducera syntes av nya proteiner involverade i neutrofil livslängd förlängning, och minskade apoptos. 10
Till skillnad från neutrofiler som är kortlivade och konstitutivt genomgår apoptos i odling, eller cytokiner / tillväxtfaktorer-aktiverade neutrofiler, som beskrivits ovan, som har förlängda livslängden, har vi nyligen identifierat en ny, liten subpopulation av neutrofiler som utvecklar spontant i långvarig standardodlings villkor från nyligen isolerade humana blod neutrofiler utan externt lägga cytokiner eller tillväxtfaktorer. 14 Dessa neutrofil-härledda celler, som inte beskrivits tidigare i litteraturen benämndes jätte fagocyter (Gφ). Den Gφ har förlängt livslängden i kultur, är de fullt utvecklade inom 5-7 dagar, och kännetecknas av unika morfologiska egenskaper, fenotypiska uttrycket och funktioner. De är kraftigt förstorade på grund av autophagocytosis av döda neutrofiler lämningar, vakuoliserade och innehåller phagolysosomes. Den Gφ uttrycker specifika neutrofila granuler markör – kluster av differentiering (CD) 66b, de azurofila granuler markörerna – CD63 och myeloperoxidas (MPO) och ytterligare neutrofiler markörer såsom CD11b, NE, CD15, de NADPH-oxidas subenheter gp91- phOx och p22- phOx och autophagy markör -LC3BII. 14, 15 Funktionellt de aktivt ta upp latexpärlor och zymosan partiklar och genererar ROS som svar på zymosan och forbol-12-myristat 13-acetat (PMA) stimulering. Intressant, till skillnad från färska neutrofiler, Gφ också intensivt uttrycka renhållare receptorer CD68 och CD36, ta upp oxiderat lågdensitetslipoprotein (oxLDL), och generera ROS som svar på stimulering med oxLDL. Dessutom Gφ saknar de monocytiska härstamning markörer CD14, CD16 och CD163 eller dendritiska markörer CD1c och CD141. Dessutom phagocytosis och autophagy och sannolikt funktionellt NADPH-oxidas är en förutsättning för deras utveckling. Detta eftersom, fagocytos-hämmaren cytochalsin B, den Autophagy inhibitorer 3-metyladenin (3-MA) och bafilomycin (BafA1) och NADPH-oxidas-hämmare – difenylen jodonium (DPI) – förhindrade deras utveckling. Dessutom, monocyter / neutrofiler samodlingar samt exponering för intermittent hypoxi hindras deras utveckling, medan neutrofila anpassning till ihållande hypoxi var uppenbar. 14,15 Deras föreslagna utvecklingen i kultur illustreras i figur 1 .Den protokoll i detta dokument beskriver steg för steg framställningen av Gφ från nyligen isolerade cirkulerande humana blod neutrofiler, deras utveckling, identifiering och vissa grundläggande egenskaper. Detta protokoll kan användas för att ytterligare undersöka och avslöja det breda spektrum och rollerna för dessa nyligen beskrivits och spännande neutrofil-härlett Gφ för att characterize deras betydelse och deras potentiella funktioner.
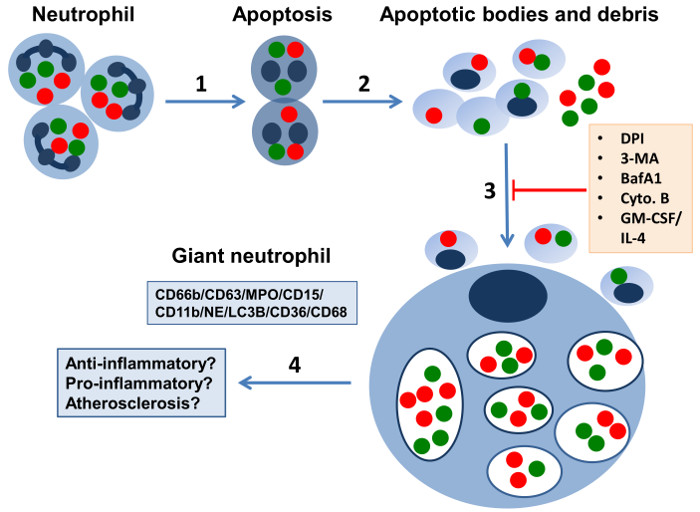
Figur 1: Schematisk representation av jätteceller utveckling 7 Dag neutrofila kulturer. Det föreslås att vid inflammatoriska ställen (1) neutrofiler genomgå apoptotisk celldöd, och (2) frigörmembran inringade fragment innehållande kärn skräp, granulat (gröna och röda prickar) och andra subcellulära komponenter som utlöser Autophagy mekanismer. (3) Jätte fagocyter (Gφ) utvecklas på lång sikt neutrofila kulturer saknar cytokiner eller tillväxtfaktorer från internalisera apoptotiska kroppar och neutrofila skräp, samtidigt som funktionella NADPH oxidase.They kännetecknas av olika neutrofilt CD66b + / CD63 + / MPO + / CD15 + / CD11b + / NE markörer, stora fagosomer omsluter granuler och cellrester, och asätare receptorer CD36 och CD68. gb1; är oftast mononukleära celler, som kan internalisera också olika partiklar och oxiderat LDL och generera ROS. Membranen i vakuoler fyllnings Gφ innehåller LC3B (markerat med mörkblå), en markör av autophagosomal membran, vilket antyder ett strikt samband mellan autophagy och jätte fagocyt bildning. Gφ utvecklar inte i medium innehållande GM-CSF / IL-4. Även inhibitorer såsom NADPH-oxidas-hämmare – difenylen jodonium (DPI), de Autophagy hämmare 3-metyladenin (3-MA) och bafilomycin (BafA1) och fagocytos hämmaren cytochalasin B (. Cyto B) avskaffa deras bildning. (4) Potentiella Gφ funktioner in vivo kan omfatta anti- eller pro-inflammatoriska egenskaper och deltagande i aterosklerotiska processer (denna siffra är baserad på våra resultat 14, 15 och ändrades från medföljande Ledare av BErton 20). Klicka här för att se en större version av denna siffra.
