העלה התירס הוא מודל אידיאלי לחקור היווצרות שדות התפתחותית במהלך המורפוגנזה, שכן יש בה גבול ברור בין הלהב לבין הנדן כי ניתן לנתיחה גנטית (איור 1 א). במהלך השלבים המוקדמים של פיתוח עלה, להקה ליניארי של תאים קטנים יותר, הלהקה preligule (PLB), subdivides העלה primordium לתחומים מראש להב וטרום-נדן. Ligule שולי דמוי auricles משולש לפתח מן PLB (איור 1 א ', ג', ד). מסך גנטי זיהה מוטציות המשבשות את גבול להב-נדן. לדוגמה, liguleless1 רצסיבי (lg1) מוטציות למחוק את ligule ו auricles 1, 2, 3, 4 (איור 1 ב). באתרו הכלאה גילתה כי תמליל lg1 מצטבר על PLB ומתפתחים ligule, מה שהופך אותו סמן מצוין לפיתוח ligule 5, 6 (איור 1E).
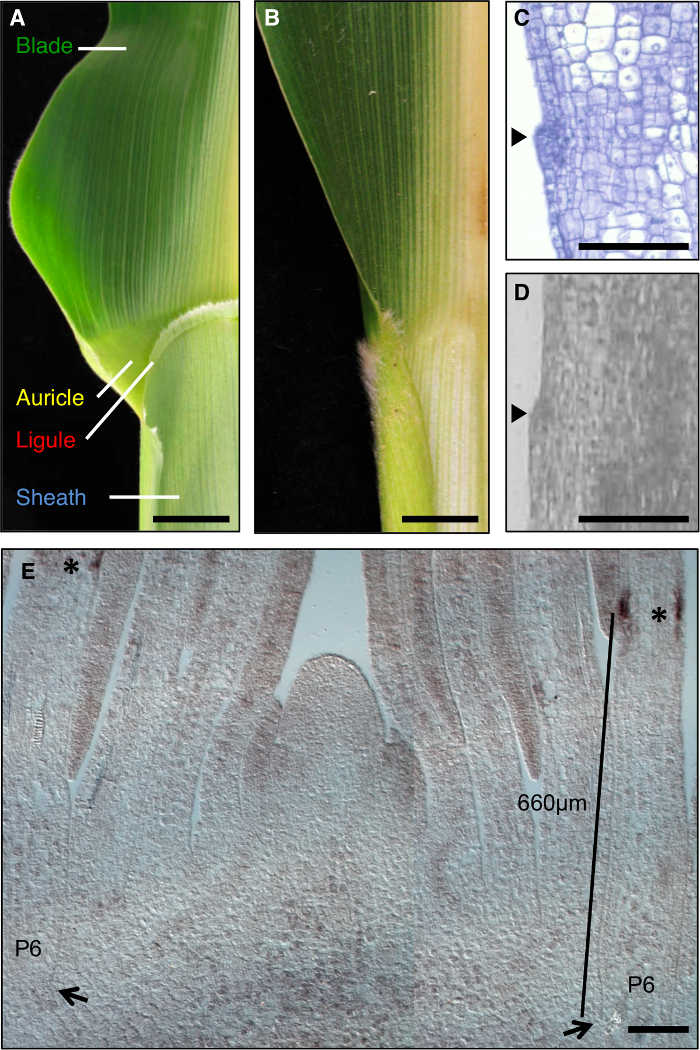
איור 1: wild-type ו liguleless1-R עלי תירס. (א) גבול Blade-נדן באזור עלה wild-type הבוגרת מראים מבני ligule ו אפרכסת. (ב) באזור הגבול Blade-נדן היעדרות מראה עלה בוגרת liguleless1-R של מבנים ligule ו אפרכסת. עלים ב A ו- B קוצצו במחצית לאורך midrib. (ג) אורך קטע דרך primordium עלה wild-type. לדוגמא תעובד ומוכתם לניתוח היסטולוגית. Ligule הייזום ניכר כבליטה הבולטת מהמטוס של העלה (ראש החץ). (ד) כת אורכיתיון דרך primordium עלה wild-type. לדוגמא תעובד עבור LM כמתואר בטקסט. ראש החץ מציין ייזום ligule. (E) lg1 הכלאה באתרו של חתך לאורך לרוחב איפקס לירות. כוכביות מצביעים הצטברות תמליל lg1 בבית PLB של primordium עלה P6. החצים מצביעים על בסיס של primordium P6. בר מציין מדידה מהבסיס של primordium אל PLB. ברי סולם ב A ו- B = 20 מ"מ. ברים בקנה מידה לספירה = 100 מיקרומטר. נתון זה יש הבדל בין התייחסות 6 (אגודה האמריקנית זכויות יוצרים של ביולוגי צמח). אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
במחקר זה, LM RNA-seq הועסק לזהות חבילה של הגנים באים לידי ביטוי באופן דיפרנציאלי (DE) בבית ביחס גבול להב-נדן לחלקים אחרים של primordium עלה ל IDE גני ntify כי הם DE ב lg1-R מוטציות ביחס אחי wild-type. LM RNA-seq היא שיטה לכימות הצטברות תמליל בתאים ספציפיים או תחומים הסלולר 7. מערכות LM לשלב לייזר מיקרוסקופ עם מצלמה דיגיטלית. רקמה מחולקת היא רכובה על שקופיות נצפית מבעד למיקרוסקופ. תוכנת LM כוללת בדרך כלל כלי ציור המאפשרים למשתמש להתוות בכל אזור שנבחר עבור microdissection. קיצוצי הליזר לאורך הקו, ואת הרקמה שנבחרו הזניקו את השקופית לתוך צינור התלוי מעל השקופית. LM מאפשר למשתמש microdissect תחומים מדויק, כולל שכבות תאים ספציפיים יחיד, גם תאי 8, 9. RNA ניתן לחלץ אז מרקמת microdissected. בהמשך לכך, מרכיב-Seq RNA מנצל רצף של הדור הבא רצף ספריות cDNA שנוצר מן RNA חילוץ 10,= "Xref"> 11.
יתרונות עיקריים של רנ"א-seq LM הם היכולת לכמת הצטברות תמליל בתחומים מוגדרים במדויק ואת היכולת גטסבי הגדול transcriptome כולו 7 זמנית. הטכניקה מתאימה במיוחד חיטוט אירועים התפתחותי מוקדם שבי את האזור של עניין הוא לעתים קרובות מיקרוסקופי. מחקרים קודמים מנוצל LM בשילוב עם הטכנולוגיה microarray ללמוד תהליכים התפתחותיים בצמחים 9, 12, 13. יש RNA-seq היתרון של לכימות תמלילים על פני טווח דינמי רחב, כוללים גנים נמוכים הביע, ומידע רצף לפני אינו נדרש 10, 11. יתר על כן, LM RNA-seq יש הפוטנציאל להדגיש גנים חשובים מבחינה התפתחותית שעשויות מוחמצות מסכי mutagenesis בשל יתירות גנטית או לערוך קטלני של האובדן-של-מוטצית פונקציה.
מבחינה התפתחותית גנים חשובים, כגון sheath1 צר (ns1) ו cotyledon2 בצורת כוס (cuc2), לעתים קרובות יש דפוסי ביטוי ספציפי אחד בלבד או מספר תאים 17, 18, 19, 20. הרבים באים לידי ביטוי רק במהלך שלבי התפתחות מוקדמים ולא באיבר הבוגר. כאשר איברים שלמים או תחומים גדולים מנותחים, תמלילי תא ספציפי אלה הם בדילול וייתכן שלא להתגלות יותר ניתוחים קונבנציונליים. על ידי התרת ניתוחים של תחומים מוגדרים במדויק, LM RNA-seq מאפשר גני הרקמות ספציפיות אלה כדי להיות מזוהים לכימות.
גורמים מכריעים בהצלחתה של הניסויים שתוארו כאן היו ניתוח היסטולוגית יסודי שהנחה מבחר הבמה המושלמת ההתפתחותית המתאימה לניתוח, ו measureme המדויקNT של תחומי רקמות תאים עבור LM. כדי להבטיח כי לדומיינים מקבילים נדגמו לכל משכפל, רקמות נאסף primordia עלה באותו שלב התפתחותי התחומים microdissected נמדדו ביחס ציוני דרך מורפולוגיים כגון ligule המתעוררים (איור 2). זה ידוע כי גנים מסוימים מתבטאים שיפוע מהקצה לבסיס של העלה. באמצעות מדידת תחומים מדויקים, וריאציה בשל דגימה ממקומות שונים לאורך הציר הפרוקסימלי-דיסטלי עלה הוחזק על (איור 3 א) מינימום. על ידי microdissecting תחומים של באותו גודל, וריאציה בשל דיפרנציאלי דילול של תמלילי תא ספציפי הופחת גם (איור 3B). סעיפים אורכים לרוחב של הקודקוד לירות שמשו כל microdissections. אלה הם חלקים כי הם בניצב לציר midrib-שוליים (איור 4). באמצעות סעיפים בלבד הכוללים את SAM מבטיח כי אזורים לרוחב מקבילים שלprimordia עלה מנותחים.
בדגימות מעובדים מחולקים עבור LM, סימן מורפולוגיים הראשון של תולדת ligule הוא בליטה בצד adaxial עקב חלוקות תא periclinal באפידרמיס adaxial (1D איור, איור 2). לעניין זה נקבע כי ligule המתעוררים יכול להיות מזוהה באופן מהימן לפי primordia עלה שלב פלסטוכרון 7. היינו מעוניינים גנים הביעו באזור ligule כולו, כולל ligule המתעוררים והתאים מייד דיסטלי שיהוו את האפרכסת. על מנת להבטיח כי בחירות רקמות מקבילות נעשו, גבשושית ligule שמשה כציון דרך מורפולוגיים מלבנים 100 מיקרומטר מרוכזים על בליטת ligule נבחרה LM (איור 2 א, 2 ב). מלבנים בגודל שווה של שבשבת טרום-נדן מראש נבחרו מאותו primordia עלה.
ניתוח של צמחים מוטנטים liguleless הציג challe שונהnge; מוטציות lg1-R לא יוצרים ligule, ולכן תכונה מורפולוגית זה לא יכול לשמש כדי לבחור את האזור עבור LM. במקום זאת, התחום של הצטברות תמליל lg1 ב primordia עלה wild-type נקבע, וכן באזור שיקיף תחום זה הוגדר. ניתוחים ראשוניים אלה בוצעו על השתילים מאותו שתילה כמו שימשו בסופו של דבר, מאז מחקרים קודמים הראו כי מיקומו של PLB משתנה בהתאם לתנאי הגידול. באתרו הכלאה ציינו כי תמלילי lg1 להצטבר primordia עלה PLB של P6 (איור 1E). בחרנו מיקרומטר תחום 400-900 מהבסיס של primordia עלה שהקיף את תחום ביטוי lg1 (מלבנים סגולים, איור 2 א) וכבש אזורים המקבילים אלה מן wild-type וצמחי lg1-R. כדי למזער וריאציה בתנאי הרקע וצמיחה גנטי כאשר משווים transcripהצטברות t ב lg1-R וצמחים בר-סוג, תוך הפרדת משפחות של מוטציות ואחי wild-type שמשה.
