A interacção entre as moléculas é a base da natureza. Assim, os cientistas em diversas áreas de pesquisa básica e aplicada tentar entender os princípios fundamentais de interacções moleculares de diferentes tipos. MicroScale termoforese (MST) permite aos cientistas realizar a caracterização rápida, precisa e de custo eficiente e de qualidade controlada de interações moleculares em solução, com uma livre escolha de buffers. Já existem mais de 1.000 publicações utilizando MST, a partir de 2016 sozinho, descrevendo diferentes tipos de análises, incluindo rastreios de bibliotecas, a ligação validações de eventos, ensaios de competição, e experimentos com múltiplos parceiros de ligação 1-8. Em geral, o MST permite o estudo dos parâmetros de ligação clássicos, tais como a afinidade de ligação (pM para mM), estequiometria, e termodinâmica, de qualquer tipo de interacção molecular. Uma grande vantagem de MST é a possibilidade de estudar eventos de ligação independentes do tamanho dos parceiros de interacção. mesmo chalinteracções entre lenging aptâmeros de ácidos nucleicos pequenos (15-30 nt) e alvos, tais como pequenas moléculas, drogas, antibióticos, ou metabolitos podem ser quantificados.
Atuais tecnologias state-of-the-art para caracterizar as interações aptâmero-alvo são ou lab-intenso e altamente complexa ou não quantificar aptâmero-pequena molécula interações 9,10. Superfície Plasmon Resonance (SPR) à base de ensaios de 11,12 e abordagens calorimétricos verdadeiramente livre de rótulo, como Calorimetria isotérmica de titulação (ITC) 13-15, eluição isocrática 16, o equilíbrio infiltração 17,18, em linha sondagem 19, distribuição do gel mudar ensaios, stopped- fluxo de fluorescência espectroscopia 20,21, anisotropia de fluorescência (FA) 22,23, uma única molécula de fluorescência de imagem 24,25, e Bio-camada de interferometria (BLI) 26 também são ou imprecisa ou incompatível com a molécula do aptâmero-small interacções. Outros principal questões de estes métodos são de baixa sensibilidade, o consumo elevado de amostras, imobilização, as limitações de transporte de massa em superfícies, e / ou restrições de buffer. Apenas algumas destas tecnologias oferecem controles integrados para efeitos de agregação e de adsorção.
MST representa uma ferramenta poderosa para os cientistas para superar esta limitação para estudar as interacções entre os aptâmeros e pequenas moléculas 27-29, bem como outros objectivos, tais como proteínas de 30-33. A tecnologia baseia-se no movimento de moléculas através de gradientes de temperatura. Este movimento dirigido, chamado "termoforese," depende do tamanho, carga, e camada de hidratação da molécula 34,35. A ligação de um ligando para a molécula irá alterar directamente, pelo menos, um destes parâmetros, o que resulta em uma mobilidade alterada thermophoretic. Ligantes com tamanhos pequenos podem não ter um impacto considerável em termos de mudança de tamanho do não ligado ao estado ligado, mas eles podem ter dr efeitos AMATIC sobre a camada de hidratação e / ou carga. As mudanças no movimento das moléculas após thermophoretic interacções com o parceiro de ligação permite a quantificação dos parâmetros de ligação de base 2,7,34,36,37.
Tal como representado na Figura 1A, o dispositivo consiste MST de um laser infravermelho focalizada sobre a amostra, dentro dos capilares de vidro utilizando a mesma óptica como para a detecção de fluorescência. O movimento thermophoretic de proteínas através da fluorescência intrínseca dos resíduos de triptofano 6 ou de um parceiro de interacção 3,8 marcado por fluorescência podem ser monitorizadas enquanto o laser estabelece um gradiente de temperatura (AT de 2-6 ° C). A diferença de temperatura, resultando em espaço, AT, conduz à depleção ou da acumulação de moléculas na zona de temperatura elevada, que pode ser quantificado pelo coeficiente Soret (S T):
g "/>
c representa quente a concentração na região aquecida, e c frio é a concentração na região fria inicial.
Como mostrado na Figura 1B, uma típica MST os resultados da experiência em um perfil de movimento MST (traço do tempo), que consiste em diferentes fases, que podem ser separadas pelos respectivos prazos. A fluorescência é medida inicial nos primeiros 5 s na ausência do gradiente de temperatura para definir a fluorescência a partir preciso e para verificar a fotobranqueamento ou photoenhancement. O salto de temperatura (T-Jump) representa a fase em que a fluorescência mudanças antes do movimento thermophoretic. Essa redução inicial na fluorescência depende de mudanças dependentes do calor de fl uorophore rendimento quântico. A fase termoforese segue, em que as reduções de fluorescência (ou aumento), devido ao movimento das moléculas de thermophoretic até que a distribuição em estado estacionário é atingido.O TJump reversa e concomitante difusão de moléculas de volta uorescent fl pode ser observado como indicado na figura 1B após o laser é desligado. De modo a aceder aos parâmetros de ligação de base, diferentes razões molares dos parceiros de interacção são analisados e comparados. Tipicamente, 16 diferentes proporções são estudadas numa experiência MST, enquanto que a molécula visível óptico é mantido constante e é fornecido com uma quantidade crescente de ligando não marcado. A interacção entre os dois parceiros de ligação induz alterações na termoforese, e, assim, na fluorescência normalizada, F norma, que é calculada como se segue:
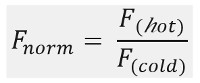
F quente e fria F representam em média intensidades fluorescência na de fi nidas pontos no tempo dos traços do MST. As afinidades de ligação (Kd ou valores de EC50) pode ser calculada por Curve encaixe (Figura 1C).
No geral, o MST é uma ferramenta poderosa para estudar interações moleculares de qualquer tipo. Este manuscrito oferece um protocolo para caracterizar a interação desafio entre a pequena molécula de trifosfato de adenosina (ATP; 0,5 kDa) e do 25-nt curta ssDNA aptâmero DH25.42 (7,9 kDa). Ao longo do manuscrito, o local de ligação do aptâmero na molécula de ATP é mapeado para baixo para o grupo de adenina do ATP.
