Analyzing Cell Surface Adhesion Remodeling in Response to Mechanical Tension Using Magnetic Beads
Summary
Cell surface adhesions are central in mechanotransduction, as they transmit mechanical tension and initiate the signaling pathways involved in tissue homeostasis and development. Here, we present a protocol for dissecting the biochemical pathways that are activated in response to tension, using ligand-coated magnetic microbeads and force application to adhesion receptors.
Abstract
Mechanosensitive cell surface adhesion complexes allow cells to sense the mechanical properties of their surroundings. Recent studies have identified both force-sensing molecules at adhesion sites, and force-dependent transcription factors that regulate lineage-specific gene expression and drive phenotypic outputs. However, the signaling networks converting mechanical tension into biochemical pathways have remained elusive. To explore the signaling pathways engaged upon mechanical tension applied to cell surface receptor, superparamagnetic microbeads can be used. Here we present a protocol for using magnetic beads to apply forces to cell surface adhesion proteins. Using this approach, it is possible to investigate not only force-dependent cytoplasmic signaling pathways by various biochemical approaches, but also adhesion remodeling by magnetic isolation of adhesion complexes attached to the ligand-coated beads. This protocol includes the preparation of ligand-coated superparamagnetic beads, and the application of define tensile forces followed by biochemical analyses. Additionally, we provide a representative sample of data demonstrating that tension applied to integrin-based adhesion triggers adhesion remodeling and alters protein tyrosine phosphorylation.
Introduction
In metazoa, mechanical tension directs tissue development and homeostasis through the regulation of a myriad of cellular processes such as proliferation, differentiation and survival 1,2. Mechanical tension can arise from the extracellular matrix or can be generated by adherent cells, which sample their extracellular environment through the actomyosin contractile machinery that pulls onto extracellular matrix and probes its rigidity through tension-sensitive molecules. In response to tension, mechanosensitive adhesion proteins undergo conformational changes that trigger complex signaling cascades. In turn, these signaling pathways orchestrate a mechanoresponse encompassing proliferation, differentiation and survival that adjusts the cellular behavior to the extracellular environment. Such processes can be settled in a short-term time period (seconds to minutes) to quickly feed back onto the loop of mechanotransduction by modifying the mechanosensitive structures. For instance, integrin-based adhesions reinforce in response to tension through Rho GTPase-mediated cytoskeletal remodeling 3,4,5. In parallel, other signaling pathways are activated over hours and days to control genetic programs that eventually impact cell fate 6. Whereas, many studies have highlighted the effect of matrix stiffness on cell determinism and disease development 1,2 , the precise molecular mechanisms of adhesion-mediated mechanotransduction still remain elusive.
Various approaches have been developed to study the effects of cell-generated forces or external forces on cell behavior, including flow systems, fluorescence resonance energy transfer (FRET)-tension sensors 7,8, compliant substrates 9, magnetic tweezers, optical tweezers 10 and atomic force microscopy (AFM) 11. Here we present a protocol using superparamagnetic beads to characterize mechanotransduction pathways in response to tensional forces applied to specific adhesion receptors. Superparamagnetic beads are particles that reversibly magnetize when placed in a magnetic field. Once coated with a ligand for a specific receptor, these beads provide a powerful tool to study the effects of extracellular force application. This method has been validated by several studies 3,5,12–17 and present the advantage to largely facilitate biochemical analysis on adherent cells. Using similar collagen-coated magnetic beads followed by biochemical analysis, early work reported an increase in protein tyrosine phosphorylation and RhoA activation in response to tension 5,18,19. The method described below has also been used with fibronectin (FN)-coated beads to characterize the signaling pathways downstream from tension applied to integrins 3. In this study, Guilluy et al. showed that tension activates RhoA through the recruitment of the two guanine nucleotide exchange factors (GEFs), LARG and GEF-H1, to integrin adhesion complexes. Since that, other studies have shown that GEF-H1 is recruited to adhesion complexes in response to cell-generated tension using different methods 20,21, demonstrating the robustness of the methodology described here. As a result, activated RhoA was shown to promote adhesion reinforcement, through cytoskeletal remodeling. This system was also used to explore tension applied to cell/cell adhesion receptors. Application of forces onto magnetic beads coated with the extracellular domain of E-cadherin induced an increase in vinculin recruitment similarly to integrin associated adhesion complexes 12. Collins and colleagues observed that application of tension to PECAM-1 promotes integrin and RhoA activation 13. Another experimental approach using magnetic beads is the study of tension applied to isolated nuclei. Using beads coated with antibodies against the nuclear envelope protein nesprin-1, nuclear envelope complexes were purified to show that they are dynamically regulated in response to mechanical tension 22. These results support the powerfulness of this method in the study of mechanotransduction pathways. Moreover, while flow or traction force systems stimulate general cellular processes, magnetic beads specifically target a cell adhesion receptor by using either receptor ligands 3 or monoclonal antibodies against cell surface receptor 13,15.
Another advantage of this method is the isolation of adhesion complexes through a straightforward ligand affinity purification procedure. It is well known that addition of ligand-coated beads to cells binds adhesion receptors and induces the recruitment of several adhesion proteins23. Further application of forces to ligand-coated magnetic beads turns these adhesion complexes into macromolecular platforms that mediates various tension-dependent signaling pathways4,24. Cell lysis followed by bead concentration using a magnet permits the isolation of the adhesion platforms. Other methods used to purify adhesion complexes have already been used in adherent cells. They combine chemical crosslinking to conserve protein-protein interactions and a cell lysis step by detergent and shear flow or sonication 20,21,25,26,27,28. The final step is the collection of the resulting ventral plasma membranes containing the adhesion complexes. Unlike these methods, magnetic beads allow a greater purification level of cell adhesion complexes by selectively targeting a specific family of adhesion receptors. Magnetic beads have already been used to purify adhesion complexes in non-adherent cells attached to ligand-coated microbeads29,30. The method described below mimics biological situations where force is applied for a short sustained period (seconds to minutes). Therefore, it provides a powerful tool for investigating both the molecular composition of purified adhesion complexes and the downstream mechanosensitive-signaling pathways.
Here we present a detailed experimental protocol for using magnetic beads to apply tensional forces to adhesion surface proteins. A permanent neodymium magnet is placed on top of the culture dish surface. The pole face of the magnet is placed at a height of 6 mm so that the force on a single 2.8 μm magnetic bead is constant (about 30-40 pN)31. The duration of tension stimulation is determined by the operator depending on the molecule of interest and its time-scale of activation. Cells are finally lysed, adhesion complexes are purified by beads separation using a magnet and biochemical analyses are processed. This protocol includes the preparation of ligand-coated superparamagnetic beads, and the application of tension through magnet followed by biochemical analyses. Additionally, we provide a representative sample of data demonstrating that tension applied to integrin-based adhesions induces adhesion remodeling and alters protein tyrosine phosphorylation.
Protocol
1. Ligand Conjugation to Magnetic Beads
Note: Ligand conjugation is performed using superparamagnetic tosyl-activated beads with a 2.8 μm diameter (stock solution concentration 108 beads/mL, 30 mg beads/mL). The following protocol is based on samples of approximately 2 x 105 cells, which correspond to MRC-5 cells grown to 80% confluency in a 60 mm tissue culture plate. Adjust the volume of beads and reagents accordingly if using plates of different sizes or cells at different confluences. Use an amount of superparamagnetic beads in order to have 2 beads per cell. Therefore, 4 x 105 beads are needed for a 60 mm plate.
- Thoroughly resuspend the beads in the original vial by vortexing at least 30 s. Aliquot 40 µL of the resuspended superparamagnetic beads (corresponding to 4 x 106 beads) to 1.5 mL microcentrifuge tube.
- Place the tube on a magnetic separation stand in order to separate the magnetic beads from the solution. Discard the supernatant, remove tube from the magnetic separation stand and resuspend the beads in 1 mL 0.1 M Na-phosphate pH 7.4.
- Repeat wash step twice with 1 mL 0.1 M Na-phosphate pH 7.4.
- After the final wash, resuspend the beads in 1 mL 0.1 M Na-Phosphate pH 7.4.
- Combine 100 μg Bovine Fibronectin (FN) or any other cell adhesion receptor ligand to the 1 mL 0.1 M Na-phosphate pH 7.4 containing beads and mix by pipetting.
- Repeat previous steps by replacing FN or any other specific ligand with BSA, poly-D-lysine or apo-transferrin for negative controls.
- Optionally, for gel analysis, take a 10 μL aliquot of ligand solution for analysis of crosslinking efficiency and mix with an appropriate volume of concentrated Laemmli Sample Buffer.
- Incubate beads with the ligand containing solution for 12-24 h at 37 °C on a rotor.
- If bead aggregates appear after the reaction has occurred, sonicate (continuous 39 W power) for no more than 10-20 s.
- Isolate the beads using magnetic separation stand, aspirate the remaining solution and add 1 mL of PBS/0.2% BSA pH 7.6 solution. Incubate for 1 h on rotor at 37 °C.
- Wash beads using a magnet twice with 1 mL PBS/0.2% BSA pH 7.6. Resuspend beads in 1 mL PBS/0.2% BSA pH 7.6.
- Sonicate beads if aggregates appear for no more than 20-30 s.
- Optionally, remove a 10 μL aliquot to analyze coupling efficiency by western blot (mix with an appropriate volume of concentrated Laemmli Sample Buffer).
- Proceed to cell assays or store beads at 4 °C for up to 1 month.
- Optionally, run a SDS-PAGE gel and stain with Coomassie blue to analyze FN crosslinking to the magnetic beads.
2. Application of Tensional Forces on the Ligand-coated Beads Bound to Adhesion Receptors on the Dorsal Surface of Cells
- Culture adherent cells on 60 mm tissue culture dish in appropriate growth medium (usually DMEM 4.5 g/L D-glucose supplemented with FCS 10%) until reaching 80% confluency.
- Prepare non-denaturing non-ionic lysis buffer (20 mM Tris HCl pH 7.6, 150 mM NaCl, 2 mM MgCl2, 0.1% NP-40) and chill microcentrifuge tubes.
- Pellet the ligand-coated beads from section 1.10 and resuspend in 1 mL PBS/0.2% BSA pH 7.6. Add 100 μL of the ligand-coated beads solution to 5 mL of warm culture medium and vortex.
- Aspirate medium in 60 mm culture dish and add 5 mL warm growth medium supplemented with beads.
- Incubate for 20 min under cell culture conditions (37 °C and 5% CO2) to allow the beads to sediment and adhere to the cells. It is crucial to not exceed 20 min since beads may be internalized by phagocytosis.
- Optionally, observe the beads under a light microscope using a 10X or 20X objective and check for bead adhesion by slightly shaking the dish to discriminate between attached and floating beads.
- While keeping the culture dish in the incubator, swap the normal dish lid for a lid with a round 38 mm neodymium magnet attached on the upper face. The magnet is held in place by two smaller 13 mm neodymium magnets positioned on the lower face of the lid. Since the magnets are very powerful, manipulate them carefully.
- Incubate cells subjected to tension for the desired time points at cell culture conditions (37 °C and 5% CO2).
- After treatment, place dish on ice and carefully aspirate the entire medium from the dish.
- Add 300 μL of cell lysis buffer, and incubate for 10 min on ice. Collect the lysate using a cell scraper and transfer to a pre-chilled 1.5 mL microcentrifuge tube.
- Pellet the magnetic beads using the magnetic separation stand and transfer the total cell lysate to a new pre-chilled 1.5 mL microcentrifuge tube. Store this fraction at -20 °C for further analysis.
- Wash the beads 3 times with 1 mL ice-cold lysis buffer. Add 50 μL Laemmli Sample buffer to the bead pellet, mix by pipetting and boil at 95 °C for 5 min using a dry block heater. This fraction contains the isolated adhesion complexes. Proceed to biochemical analysis or store at -20 °C.
- From the total cell lysate obtained after beads separation (about 300 μL), remove a 50 μL aliquot for western blot analysis.
NOTE: The 250 μL left can be used for further biochemical studies such as protein-protein interaction studies (immunoprecipitation, GST pull-down assays) or GTPase activity experiments (changing the lysis buffer may be necessary depending on the GTPase of interest).
Representative Results
The schematic of the technique is illustrated in Figure 1a. Following ligand conjugation, magnetic beads are incubated with cells for 20 min, and then a permanent magnet is used to apply tensile forces of about 30-40 pN for various amount of time. Figure 1b shows 2.8 µm FN-coated magnetic beads bound to MRC5 cell adhesion receptors.
The wash steps of superparamagnetic beads after cell lysis are crucial and determine the degree of purification. A minimum of three washes is recommended. GADPH immunoblots with long exposures can be useful to test the purity of adhesion complexes (Figure 2a).
FN-coated beads were used to investigate the mechanotransduction processes that occur over time at the adhesion complexes in response to tension. After magnetic separation of the adhesion complex fraction, the lysate and the adhesion complex fraction were analyzed by western blot. As expected, we observed talin, vinculin and paxillin, but not GAPDH in the adhesion complexes fraction even in absence of mechanical stimulation (Figure 2b). Consistent with previous reports32,33, tension triggered vinculin recruitment to adhesion complexes. While tension did not affect paxillin recruitment to adhesion complexes, its phosphorylation on tyrosine 31 was enhanced in response to mechanical tension both in the total cell lysate and in the adhesion complex fraction.
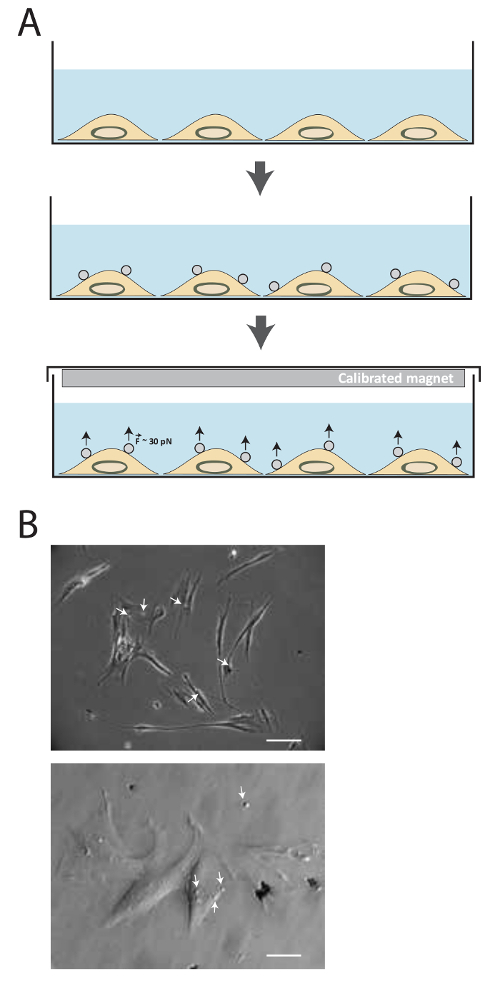
Figure 1. Description of the method. (A) Schematic illustration of the technique. Cells are firstly cultured in medium culture until the desired confluency is reached. Then, superparamagnetic beads are added for 15-20 min. Tensional forces about 30-40 pN are then applied using the calibrated magnet for different amount of time. (B) Fibronectin-coated superparamagnetic beads bind to cell adhesion receptors. Cells are imaged by phase contrast in transmitted-light microscopy 15 min after adding the superparamagnetic beads (white arrow) in medium. (Top image: Scale bar= 25 µm, bottom image: Scale bar = 100 µm) Please click here to view a larger version of this figure.
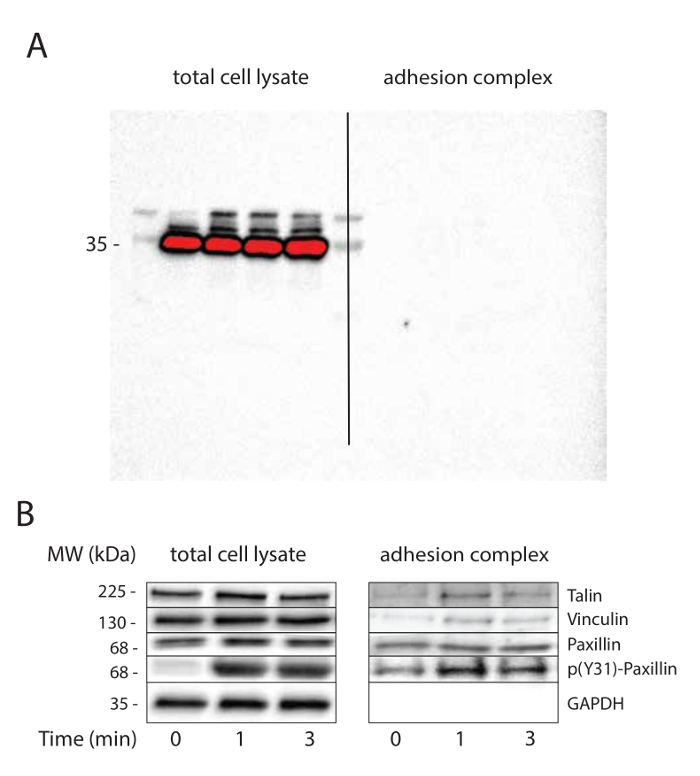
Figure 2. Mechanical tension induces adhesion maturation. (A) Purification of adhesion complexes. GAPDH immunoblotting is used as loading control for total cell lysate and to check the purity of adhesion complexes. The nitrocellulose membrane has been imaged using a long exposure to demonstrate the absence of signal in adhesion complex fraction. (B) Tension induces adhesion maturation. Loading control (GAPDH) and candidates (vinculin, talin and paxillin) known to be recruited or phosphorylated at adhesion complexes in response to mechanical tension are immunoblotted. This experiment has been performed on MRC5 cells. Please click here to view a larger version of this figure.
Discussion
The method described here constitutes a straightforward approach to apply tension to cell surface adhesion receptors and allow their subsequent purification. However, some steps are critical to perform efficient adhesion purification and potential optimization can be done depending on the targeted adhesion receptors. We present potential issues the user may encounter below.
We used 2.8 μm diameter magnetic beads but larger beads can be used such as 4.5 μm diameter. However, bead diameter should be restricted to 2-5 μm since phagocytosis might occur more quickly during 30-60 min incubation and larger beads have stronger adhesion so that bead displacement would be restricted under magnetic field34,35. Therefore, it is important to limit incubation time to short periods and use the proper bead size. The number of beads incubated per cell will impact the amount of force experience by a single cell. While one wants to efficiently stimulate cells with tension, too many beads per cell can lead to activation of irrelevant signaling pathways. We usually incubate an average of two beads per cell for FN-coated beads, but this quantity can be adjusted [1 to 5 beads per cell] depending on the cell type and the cell surface receptor. Regarding the magnet, the method described here used a magnet for which the resulting force on a 2.8 μm has been measured (about 30-40 pN by measuring the displacement of magnetic beads in undiluted glycerol, a Newtonian liquid with known viscosity31, although it is possible to use larger magnets to apply larger amount of force if necessary. It is important to note that these magnets are extremely powerful and to manipulate them carefully.
After 12-24 h incubation with ECM ligand (step 1.7) as well as after 1 h incubation (step 1.9) with PBS/0.2% BSA pH 7.6 buffer, the magnetic microbeads might form aggregates. It is then critical to separate beads as much as possible to respect the ratio of 2 beads per cell. However, it is still acceptable to have a 5:1 ratio. Beads separation can be achieved either by mixing and pipetting or by using sonication, although for a short period (20-30 s).
Various proteins can be conjugated to the beads, including integrin ligand (FN, collagen), recombinant proteins or antibodies targeting specific cell surface receptors. The attachment of ligand-coated magnetic beads to the dorsal cell surface might be weakened by the low coupling efficiency of the ligand onto the beads. It is recommended to check for ligand binding to the beads by collecting an aliquot of the diluted ligand solution before adding to the beads and an aliquot of beads at the end of the coupling process. These aliquots can be processed to SDS-PAGE analysis and Coomassie blue staining.
If no changes in signaling pathways or expected mechanosensing processes are detected, it is important to consider different possibilities. One way to identify the issue is to modulate the duration of the experiment with the magnet. It is well known that the velocity of cellular processes varies depending on the cell type used. Another option would be to test known tension-sensitive molecules and check for their post translational modifications by western blotting (for instance paxillin phosphorylation or FAK phosphorylation can be analyzed). The amount of force may be also critical and using a thicker magnet (same grade N52) or larger beads can be an option. Additionally, ligand/receptor binding efficiency can be explored by analyzing adhesion complex and look for cell surface receptors (such as cadherin or integrin). The cell confluency can also affect the cellular response to tension. As it has been observed that cell/cell adhesion can affect cell behavior and cystokeletal prestress, it is important to note that the confluency may affect the cellular response to tension. Even though we recommend 80% confluency for application of tension to integrin-based adhesions, different conditions may be tested in order to optimize the experimental system.
Although this method provides a powerful tool to decipher the tension sensitive adhesome as well as the associated signaling pathways, it also had some limitations. First, since the main concern using such small size magnetic beads is the risk of their internalization by the cells, this method cannot be used for studying long-term mechanosensitive signaling cellular responses that modify cell fate such as differentiation and proliferation. Another limitation lies in the way of how forces are applied to the layer of cells in the culture plate. Indeed, since the magnetic field is always higher at the periphery of the magnet, forces could vary at the surface of the culture plate with a decreasing gradient of cell stretching from the periphery to the center and might lead to heterogeneous cellular responses.
The procedure described here constitutes a straightforward and cost effective method that allows studying mechanosensitive cellular pathways as well as investigating the molecular composition of adhesion complexes subjected to tension. Other approaches, such as stretching device that apply cyclic strain to cells, lead to the application of tension to all cell surface receptors interacting with the extracellular matrix. The method described here has the advantage of enabling force stimulation of a specific subset of cell surface receptors and a large variety of ligands can be used, such as integrin ligands or antibodies targeting cell surface receptors, allowing the study of many distinct mechanosensitive systems. Another advantage of this method is that it leads to the purification of the proteins complexes that experienced tension and to work on loadbearing elements which are known to play a central role in mechanotransduction36. The purified adhesion complex can also be used for various biochemical approaches14, such as kinase assays to investigate kinase activity in response to tension, or actin polymerization assay. Additionally, this experimental system can be coupled with magnetic tweezers to explore the cellular mechanical response and to correlate this response with the identified signaling pathways. Interestingly, magnetoplasmonic nanoparticles have been used recently to mechanically load Notch and E-cadherin with precise control in time and space37. This recent development may help exploring mechanosensitive signaling pathways with different spatial, temporal and mechanical inputs.
Divulgazioni
The authors have nothing to disclose.
Acknowledgements
C.G. is supported by grants from the Agence National de la Recherche (ANR-13-JSV1-0008), from the European Union Seventh Framework Programme (Marie Curie Career Integration n˚8304162) and from European Research Council (ERC) under European Union's Horizon 2020 research and innovation program (ERC Starting Grant n˚639300).
Materials
| Neodymium magnets (on the upper face of 60 mm dish) | K&J Magnetics, Inc | DX88-N52 | grade N52 dimension: 1 1/2" dia. x 1/2" thick |
| Neodymium magnets (on the lower face of 60 mm dish) | K&J Magnetics, Inc | D84PC-BLK | grade N42 dimension: 1/2" dia. x 1/4" thick Black Plastic Coated |
| Dynabeads M280 Tosylactivated | Thermofisher | 14203 | superparamagnetic beads |
| DynaMag-2 Magnet | Thermofisher | 12321D | |
| Fibronectin | Sigma-Aldrich | F1141-5MG | Fibronectin from bovine plasma |
| Poly-D-Lysine | Sigma-Aldrich | P7280-5MG | |
| Apo-Transferrin | Sigma-Aldrich | T1428-50MG | Bovine Apo-Transferrin |
| Bovine serum albumin | Sigma-Aldrich | A7906-500G | |
| DMEM high glucose, GlutaMAX supplement, pyruvate | Life Technologies | 31966-021 | DMEM+GlutaMAX-I 500 ml |
| 60*15 mm culture dish | Falcon | 353004 |
Riferimenti
- Discher, D. E., Janmey, P., Wang, Y. -. L. . Tissue cells feel and respond to the stiffness of their substrate. 310 (5751), 1139-1143 (2005).
- DuFort, C. C., Paszek, M. J., Weaver, V. M. Balancing forces: architectural control of mechanotransduction. Nat Rev Mol Cell Biol. 12 (5), 308-319 (2011).
- Guilluy, C., et al. The Rho GEFs LARG and GEF-H1 regulate the mechanical response to force on integrins. Nat Cell Biol. 13 (6), 722-727 (2011).
- Matthews, B. D., Overby, D. R., Mannix, R., Ingber, D. E. Cellular adaptation to mechanical stress: role of integrins, Rho, cytoskeletal tension and mechanosensitive ion channels. J Cell Sci. 119 (3), 508-518 (2006).
- Zhao, X. -. H., et al. Force activates smooth muscle alpha-actin promoter activity through the Rho signaling pathway. J Cell Sci. 120 (Pt 10), 1801-1809 (2007).
- Engler, A. J., Sen, S., Sweeney, H. L., Discher, D. E. Matrix elasticity directs stem cell lineage specification. Cell. 126 (4), 677-689 (2006).
- Austen, K., Kluger, C., Freikamp, A., Chrostek-Grashoff, A., Grashoff, C. Generation and analysis of biosensors to measure mechanical forces within cells. Meth Mol Biol. 1066, 169-184 (2013).
- Grashoff, C., et al. Measuring mechanical tension across vinculin reveals regulation of focal adhesion dynamics. Nature. 466 (7303), 263-266 (2010).
- Pelham, R. J., Wang, Y. l. . Cell locomotion and focal adhesions are regulated by substrate flexibility. Proc Natl Acad Sci USA. 94 (25), 13661-13665 (1997).
- Choquet, D., Felsenfeld, D. P., Sheetz, M. P. Extracellular matrix rigidity causes strengthening of integrin-cytoskeleton linkages. Cell. 88 (1), 39-48 (1997).
- Chaudhuri, O., Parekh, S. H., Lam, W. A., Fletcher, D. A. Combined atomic force microscopy and side-view optical imaging for mechanical studies of cells. Nat Meth. 6 (5), 383-387 (2009).
- Bays, J. L., et al. Vinculin phosphorylation differentially regulates mechanotransduction at cell-cell and cell-matrix adhesions. J Cell Biol. 205 (2), 251-263 (2014).
- Collins, C., et al. Localized tensional forces on PECAM-1 elicit a global mechanotransduction response via the integrin-RhoA pathway. Curr Biol. 22 (22), 2087-2094 (2012).
- Gordon, W. R., et al. Mechanical Allostery: Evidence for a Force Requirement in the Proteolytic Activation of Notch. Dev Cell. 33 (6), 729-736 (2015).
- Lessey-Morillon, E. C., et al. The RhoA guanine nucleotide exchange factor, LARG, mediates ICAM-1-dependent mechanotransduction in endothelial cells to stimulate transendothelial migration. J Immunol. 192 (7), 3390-3398 (2014).
- Osborne, L. D., et al. TGF-β regulates LARG and GEF-H1 during EMT to affect stiffening response to force and cell invasion. Mol Biol Cell. 25 (22), 3528-3540 (2014).
- Scott, D. W., Tolbert, C. E., Burridge, K. Tension on JAM-A activates RhoA via GEF-H1 and p115 RhoGEF. Mol Biol Cell. 27 (9), 1420-1430 (2016).
- Glogauer, M., Ferrier, J., McCulloch, C. A. Magnetic fields applied to collagen-coated ferric oxide beads induce stretch-activated Ca2+ flux in fibroblasts. Am J Physiol – Cell Physiol. 269 (5), C1093-C1104 (1995).
- Glogauer, M., et al. Calcium ions and tyrosine phosphorylation interact coordinately with actin to regulate cytoprotective responses to stretching. J Cell Sci. 110 (Pt 1), 11-21 (1997).
- Kuo, J. -. C., Han, X., Hsiao, C. -. T., Yates, J. R., Waterman, C. M. Analysis of the myosin-II-responsive focal adhesion proteome reveals a role for β-Pix in negative regulation of focal adhesion maturation. Nat Cell Biol. 13 (4), 383-393 (2011).
- Schiller, H. B., et al. β1- and αv-class integrins cooperate to regulate myosin II during rigidity sensing of fibronectin-based microenvironments. Nat Cell Biol. 15 (6), 625-636 (2013).
- Guilluy, C., et al. Isolated nuclei adapt to force and reveal a mechanotransduction pathway in the nucleus. Nat Cell Biol. 16 (4), 376-381 (2014).
- Plopper, G. E., McNamee, H. P., Dike, L. E., Bojanowski, K., Ingber, D. E. Convergence of integrin and growth factor receptor signaling pathways within the focal adhesion complex. Mol Biol Cell. 6 (10), 1349-1365 (1995).
- Roca-Cusachs, P., Gauthier, N. C., Del Rio, ., A, M. P., Sheetz, Clustering of alpha(5)beta(1) integrins determines adhesion strength whereas alpha(v)beta(3) and talin enable mechanotransduction. Proc Natl Acad Sci USA. 106 (38), 16245-16250 (2009).
- Ajeian, J. N., et al. Proteomic analysis of integrin-associated complexes from mesenchymal stem cells. Proteomics Clin Appl. 10 (1), 51-57 (2016).
- Horton, E. R., Astudillo, P., Humphries, M. J., Humphries, J. D. Mechanosensitivity of integrin adhesion complexes: Role of the consensus adhesome. Exp Cell Res. , (2015).
- Jones, M. C., et al. Isolation of integrin-based adhesion complexes. Curr Protoc Cell Biol. 66, 9.8.1-9.8.15 (2015).
- Ng, D. H. J., Humphries, J. D., Byron, A., Millon-Frémillon, A., Humphries, M. J. Microtubule-dependent modulation of adhesion complex composition. PloS One. 9 (12), e115213 (2014).
- Byron, A., Humphries, J. D., Bass, M. D., Knight, D., Humphries, M. J. Proteomic analysis of integrin adhesion complexes. Sci Sign. 4 (167), pt2 (2011).
- Byron, A., Humphries, J. D., Craig, S. E., Knight, D., Humphries, M. J. Proteomic analysis of α4β1 integrin adhesion complexes reveals α-subunit-dependent protein recruitment. Proteomics. 12 (13), 2107-2114 (2012).
- Marjoram, R. J., Guilluy, C., Burridge, K. Using magnets and magnetic beads to dissect signaling pathways activated by mechanical tension applied to cells. Methods. , (2015).
- Pasapera, A. M., Schneider, I. C., Rericha, E., Schlaepfer, D. D., Waterman, C. M. Myosin II activity regulates vinculin recruitment to focal adhesions through FAK-mediated paxillin phosphorylation. J Cell Biol. 188 (6), 877-890 (2010).
- Sawada, Y., Sheetz, M. P. Force transduction by Triton cytoskeletons. J Cell Biol. 156 (4), 609-615 (2002).
- Grinnell, F., Geiger, B. Interaction of fibronectin-coated beads with attached and spread fibroblasts. Binding, phagocytosis, and cytoskeletal reorganization. Exp Cell Res. 162 (2), 449-461 (1986).
- Schroeder, F., Kinden, D. A. Measurement of phagocytosis using fluorescent latex beads. J Biochem Biophys Meth. 8 (1), 15-27 (1983).
- Hoffman, B. D., Grashoff, C., Schwartz, M. A. Dynamic molecular processes mediate cellular mechanotransduction. Nature. 475 (7356), 316-323 (2011).
- Seo, D., et al. A Mechanogenetic Toolkit for Interrogating Cell Signaling in Space and Time. Cell. 165 (6), 1507-1518 (2016).
