Memeli genomları yaklaşık 20,000 protein kodlama genleri ihtiva olduğu tahmin edilmektedir. Alternatif yapıştırma ve translasyon sonrası modifikasyonlar, protein repertuarı arttırır. Proteinler, modüler bir yapıya 1 ve genellikle farklı protein kompleksleri halinde kendi işe ve birden fazla hücresel süreçleri 2 katılımlarını izin birden fazla etkileşim bölgeleri içerir. Bir örnek p120ctn adlandırılan, çok fonksiyonlu bir proteindir. p120ctn Ctnnd1 geni tarafından kodlanan ve bir N-terminal ve C-terminal bölgesi tarafından kuşatılır geniş bir merkezi armadillonun tekrar alanı oluşur. p120ctn bir armadillonun alan, hücre-hücre adhezyonu ile ilgili olan klasik kaderinler, yüksek oranda korunan bir jukstamembran alanı bağlanan, ancak aynı zamanda transkripsiyonel represör Kaiso bağlanır. p120ctn N-terminal alanı, farklı kinaz, fosfataz, küçük RhoGTPases ve mikrotübüle bağlı p ile etkileşimeroteinler 3. İlginç bir şekilde, alternatif bağlantısının sonucu olarak, p120ctn izoformları dört alternatif bir başlangıç kodonu 4 elde edilebilir. başlatma kodonu de en çok 5' çevrilmiş ve tam uzunlukta, N-terminal segmenti ihtiva gibi p120ctn izoform 1A, en uzun. p120ctn 3 ve 4, bu N-terminal segmenti, kısmen ve tamamen sırasıyla silinir izoformları. farklı hücresel fonksiyonları protein (ya da protein izoformlarının) ve bunların bölgesinin tam rolünü anlamak bir sorun olmaya devam etmektedir.
mESCs gen hedefleme karşılık gelen genin genetik silerek bir proteinin fonksiyonunun çalışma sağlar ve yaygın olarak, gelişimsel olarak önemli ve hastalığa, ilgili genlerin ve yolların belirlenmesi katkıda bulunmuştur. Ters genetik Bu atılım nedeniyle homolog yeniden 5'e Mesc izolasyon ve gen hedefleme alanlarındaki gelişmelerin sonucuydu </s> Yukarı. Homolog rekombinasyon, DNA fragmanları, çift sarmallı (ds) DNA kırılmalarının sonra, iki benzer veya özdeş nükleik yarımları arasında değiş tokuş edildiği bir süreçtir. dsDNA sonları seyrek olduğu için Normalde İK verimsizdir. Son zamanlarda, hedefleme homoloji yönlendirilmiş gen etkinliği site-spesifik nükleazlar 6, 7 kullanılarak arttırılabilir, ancak ne yazık ki bu hedef dışı etkiler 8 yatkındır. Daha güvenilir bir teknik Gen hedefleme sağlamak için Cre / loxP veya FLPe / FRT gibi site-spesifik rekombinasyon sistemleri dayanmaktadır RMCE vardır. LoxP ve Frt dizisi bakteriyofaj P1 bulunurlar ve Saccharomyces cerevisiae ve sırasıyla site yönünü belirleyen bir asimetrik 8 bp sekansı da dahil olmak üzere, 34 bp oluşur. Diğer yandan, oryantasyonu, örneğin, bir DNA bölümü içindeki iki loxP floxed DNA kesilmiş olur belirlemek veya i olacaktırKre-aracılı rekombinasyon 9 üzerine nversed. iki site farklı kromozom üzerinde bulunan Üstelik, eğer Kre da translokasyon neden olabilir. RMCE çapraz reaksiyona girmez ve bir genomik gömülü heterospesifik rekombinasyon yararlanır. Aynı heterospesifik bölgeleri tarafından takviye eden bir DNA fragmanı ihtiva eden bir Verici plazmit varlığında, rekombinaz için çift eş zamanlı translokasyon (Şekil 1) RMCE uyumlu genomik bu DNA fragmanı ekleyecektir. Burada, sadece doğru RMCE hedefli klonlar geri gelen vektör üzerinde bir promotere ilaç direnci sayesinde işleyebilen bir "tuzak" promotör az Neomisin direnç geni yerleştirme hücreleri R26 genomunda bulunan (NeoR) (Şekil 1) 10, 11. Bu <% 100 11 ile çok yakındır çok yüksek hedefleme verimliliği, sonuçlanır/ sup> 12. Sonuç olarak, RMCE bazlı bir hedefleme son derece verimlidir ve yapı-işlev çalışmaları için kullanılabilir; Bununla birlikte, bu, önceden tasarlanmış bir genomik lokus gerektirir.
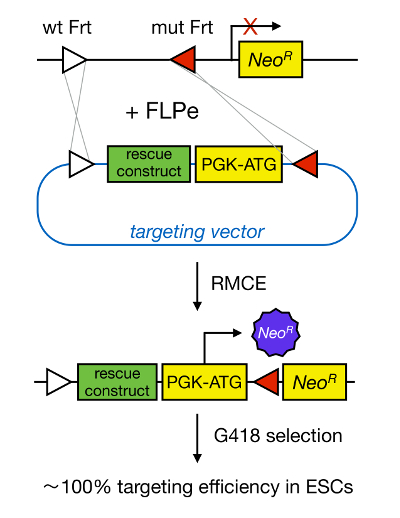
RMCE aracılı Hedefleme Şekil 1 şematik gösterimi. Her iki heterospesifik FRT bölgeleri (beyaz ve kırmızı üçgenlerle gösterilir) liman halinde RMCE eden tanımlı bir genomik lokus gelen bir hedefleme vektörü DNA segmentlerinin alışverişine izin verir. Buna ek olarak, işlenmiş genomik bölgenin bir promotörsüz ve tepe bölümü kesik neomisin direnci (NeoR) genini içerir. Gelen DNA fragmanı kodonu bir promotörü ve başlangıç olarak, yalnızca doğru rekombinasyon olayları, yüksek hedefleme verimi ile, neomisin direncini yeniden. T daha büyük bir versiyonunu görmek için buraya tıklayınOnun rakam.
mESCs içinde Genom mühendislik RMCE uyumlu farelerin üretilmesine imkan tanır. 1981 yılında, iki grup blastosist iç hücre kütlesinin (ICM) pluripotent hücreler yakalama ve kültür 13, 14 onları korumuştur. mESCs germ hücresi soyundan dahil embriyonik ve yetişkin hücrelerinin her türlü içine kendini yenileme ve farklılaşma yeteneğine sahiptir. Bu nedenle, mESCs içinde gen hedefleme konstitütif veya (Cre / LoxP sistemi kullanılarak) koşullu KO fareleri geliştirilmesi yoluyla ters genetik çalışmalar sağlar. Ancak, fare ES hücreleri izole etmek için klasik yol çok verimsizdir. Birkaç büyük gelişmeler büyük ölçüde SR ve fetal sığır serumu (FBS) 16 içeren Mesc ortam arasında değişen, 15 orta tanımlanmış serum yerine (SR) kullanımı da dahil olmak üzere, türetme Mesc hatları için başarı oranını artırmıştır ve İlaçlarla kullanımı adresBu tür pluripotin veya 2i 17 mantıksal bileşikler. Pluripotin, küçük sentetik bir molekül, lösemi inhibitör faktörü (LİF) ve fare embriyonik fibroblastlar (MEF'ler) 18 yokluğunda farklılaşmamış bir halde mESCs arasında yayılmasını sağlar. Son olarak, mESCs SR / FBS ortamı değiştirme kuralı, 20, LİF ve 19 pluripotin birleştirildiğinde (% 100'e yakın), çok yüksek bir verimle izole edilebileceği gösterilmiştir. Bu protokoller, daha sonra yapı-işlev çalışmaları için kullanılabilir RMCE uyumlu KO mESCs etkili izolasyonunu sağlar.
Bu çalışma, spesifik bir hücre süreçlerinden sorumludur bir protein içinde anahtar alanları ya da kalıntılarını tanımlamak üzere sağlayan bir yöntemi tarif etmektedir. Bu amaçla, verimli Mesc izolasyonunu sağlayan gelişmiş teknolojilerin, hedefleme vektörü montaj ve Mesc hedeflemenin bir boru hattı oluşturmak oldud. Protein izoform alan mutantlar ve devamındaki efektörlere böyle büyük paneller KO mESCs sokulabilir ve in vitro KO fenotipi kurtarmak için kabiliyetleri açısından değerlendirilebilir gibi.
