The dot assay presented here (Figure 1) was performed using invasive, lung colony forming breast cancer cells (MCF10CA1a) as a model system. The dot assay yields highly reproducible cell dots even in the hand of beginners, making it a convenient and easy to execute assay (Figure 1D). The dot assay combined with HE staining, or time-lapse imaging and subsequent particle image velocimetry (PIV) allows the study of various migration parameters including displacement of epithelial edges, cell speed, and cell directionality as cells at the edges of an epithelial sheet migrate into a cell-free area. Initially, phase contrast imaging of cell dots revealed that invasive, lung colony forming breast cancer cells (MCF10CA1a) maintain a mesenchymal phenotype after stimulation with EGF. However, single cells leaving the sheet are observed more frequently in EGF-stimulated cell dots (Figure 2, arrows). The response of MCF10CA1a cells to EGF was further assessed by HE staining of cultures that were stimulated with EGF for 4 days. Indeed, EGF-stimulation resulted in an increased colony diameter (Figure 3). These observations indicate that migration of EGF stimulated cells might be altered.
Therefore, the dot assay was next utilized to perform a more detailed analysis of the dynamic migration phenotype of MCF10CA1a cell sheets. Cells at the sheet edges were imaged for 9 h following stimulation of cells with EGF (Video 1 and Video 2) and images were then analyzed by PIV in Matlab4,13,14. PIV revealed that EGF increases cell speed (Figure 4A) to 0.63 µm/min from 0.45 µm/min in control cells. At the same time, EGF increased directionality of cells, and this is indicated by a reduction of the variability of cell directionality (angular spread) from 0.95 in control cultures to 0.79 in EGF-stimulated cultures (Figure 4B). EGF increased the radial displacement of the epithelial edge over 9 h from 257 µm to 356 µm (Figure 4), as was also observed by HE staining after 4 days (Figure 3). Thus, the use of the dot assay in combination with different subsequent assays shows that EGF alters migration at the edges of MCF10CA1a cells sheets such that the cell speed and directionality are increased, resulting in an increased colony radius.
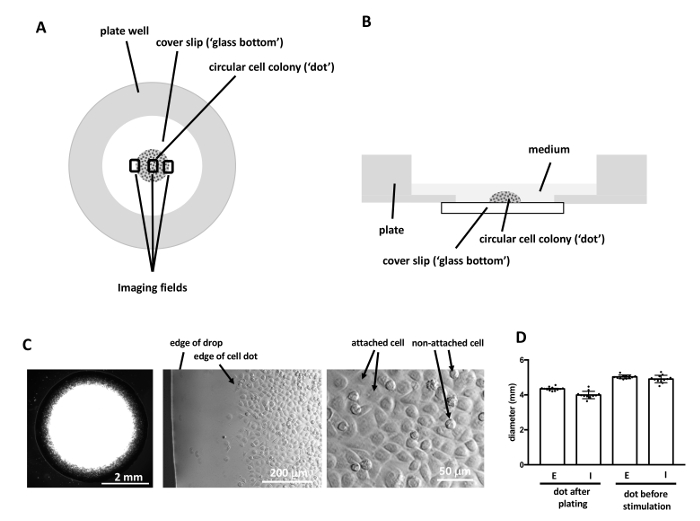
Figure 1: Imaging the dot assay. (A, B) Cells are plated as a circular colony in the center of glass bottom plate wells. For migration analysis, images are taken at two opposing edges as well as the center of the colony (A). (C) Dark field image of cell dot during plating (left), and in higher magnification (phase contrast) showing the edge of the drop and the edge of the cell dot after cells attached to the plate (center). Attached cells are flat and polygonal, while non-attached cells are round (right). (D)The diameter of dots is highly reproducible. Dots were imaged immediately after plating and before cells were stimulated and the diameter determined using ImageJ. The dot size is highly reproducible within one experiment as well as between different users and little training is required to gain proficiency in plating the cell dots (E = more than 10 years of experience in tissue culture, I = less than 4 weeks of experience in tissue culture). n = 12 dots per group. Please click here to view a larger version of this figure.
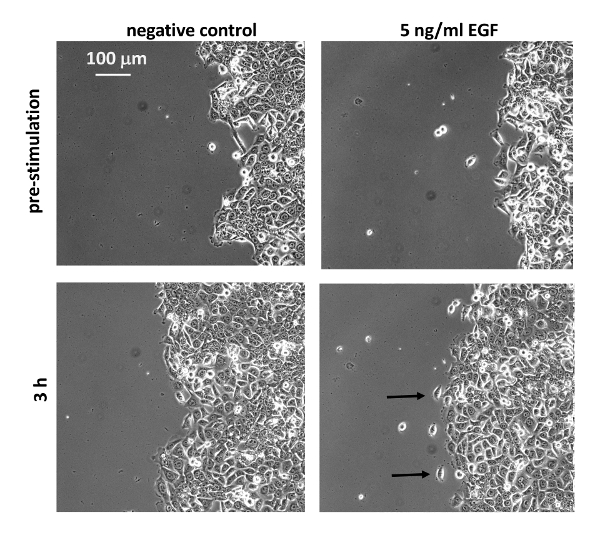
Figure 2: EGF modulates the morphological phenotype of MCF10CA1a cells. MCF10CA1a cells were starved for 3 h in DMEM/F12 supplemented with 0.1% horse serum before they were stimulated with EGF (5 ng/mL). At the edge of EGF-stimulated colonies single cells tend to migrate out of the sheet (arrows). Please click here to view a larger version of this figure.
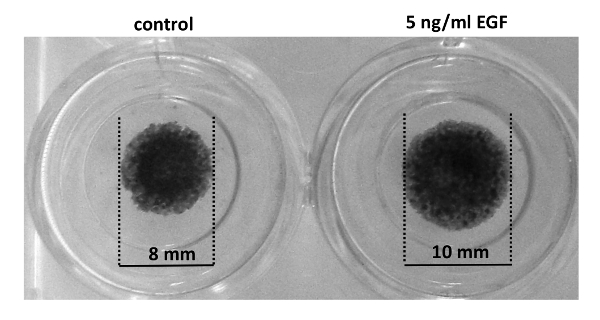
Figure 3: EGF increases net growth of cell colonies. MCF10CA1a cells were starved for 3 h in DMEM/F12 medium supplemented with 0.1% horse serum before they were stimulated with EGF (5 ng/mL). 4 days later, cells were ethanol fixed and stained with Hematoxylin & Eosin. EGF-stimulated dots have a higher diameter after 4 days. Please click here to view a larger version of this figure.
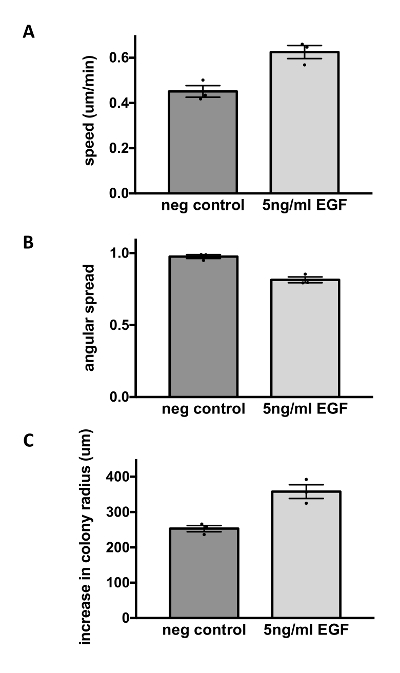
Figure 4: EGF modulates the migratory phenotype of MCF10CA1a cells. MCF10CA1a cells were starved for 3 h in DMEM/F12 medium supplemented with 0.1% horse serum before they were stimulated with EGF (5 ng/mL). Cells were imaged every 3 min for 9 h. Subsequent particle image velocimetry analysis was performed in Matlab13. Speed values represent the average speed over time for each field. Angular spread of velocity vectors was calculated as:
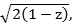 where
where 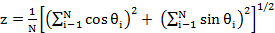 , and ranges from 0 (high directionality) to
, and ranges from 0 (high directionality) to 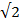 (low directionality). A decrease in angular spread is considered an increase in directionality. Radial displacement was calculated by subtracting the colony radius at t = 0 h from the colony radius at t = 9 h. EGF-stimulation of MCF10CA1a cell dots resulted in increased cell speed (A; upper panel), decreased angular spread or increased directionality (B; middle panel), and increased colony radius (C; bottom panel). Data represent mean ± SEM of n = 3 experiments. Please click here to view a larger version of this figure.
(low directionality). A decrease in angular spread is considered an increase in directionality. Radial displacement was calculated by subtracting the colony radius at t = 0 h from the colony radius at t = 9 h. EGF-stimulation of MCF10CA1a cell dots resulted in increased cell speed (A; upper panel), decreased angular spread or increased directionality (B; middle panel), and increased colony radius (C; bottom panel). Data represent mean ± SEM of n = 3 experiments. Please click here to view a larger version of this figure.
Video 1: Time-lapse imaging of unstimulated MCF10CA1a cells. Images were taken every 3 min for 9 h. Please click here to view this video. (Right-click to download.)
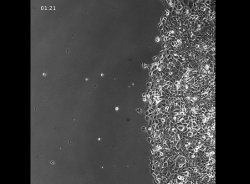
Video 2: Time-lapse imaging of MCF10CA1a cells stimulated with EGF (5 ng/mL). Images were taken every 3 min for 9 h. Please click here to view this video. (Right-click to download.)
